| Головна |
| ЗМІСТ | »» |
|---|
В результаті вивчення даного розділу студент повинен:
знати
вміти
володіти
- навичками системного аналізу будови НС в еволюційному ряду тварин.
Вивчення філогенезу НС має велике значення для морфології. Шляхом вивчення розвитку органу в системі живих організмів легше зрозуміти особливості і зміст його анатомічної структури, і за цієї умови анатомія втрачає статичний характер і перестає бути простим описом структурних співвідношень, хоча б і дуже складних.
Різноманітні зв'язку із зовнішнім середовищем властиві будь-якої живої матерії. На відміну від рослин, тварин необхідно мати можливість активно переміщатися в навколишньому середовищі, в першу чергу - для добування їжі. В процесі еволюції тварин рухова функція постійно вдосконалюється.
У одноклітинних тварин існує три основних способи руху - амебоідное, за допомогою джгутиків, за допомогою війок. Для узгодженого руху війок у інфузорій (війчастих найпростіших) виникають специфічні фібрили, здатні проводити збудження краще, ніж гіалоплазма. У багатоклітинних тварин зберігаються всі ці типи руху в клітинах, що виконують іншу функцію. Наприклад, амебоідное рух - у фагоцитів крові і мікрогліальних клітин, джгутики - у сперматозоїдів, ресничний миготливийепітелій - в бронхах.
З цього видно, що при зміні рівня розвитку на більш високий в ньому зберігаються стародавні форми організації, якщо вони можуть виконувати якусь більш приватну функцію.
Виникнення НС пов'язано з появою багатоклітинних тварин. Щоб всі клітини організму працювали злагоджено, між ними повинна існувати система зв'язків. За відсутності НС передача збудження для узгоджених дій можлива за рахунок елементів мембрани і цитоплазми, але така передача, по-перше, дуже повільна і, по-друге, не може поширюватися на великі відстані - хвиля збудження загасає. Поява НС, що надає можливість передавати інформацію швидко і на великі відстані (ПД поширюється по мембрані нейрона без загасання, див. Пункт 2.7), дає організму великі переваги в адаптації до навколишнього середовища. Якщо порівняти два типи безхребетних - губок і кишковопорожнинних, що мають багато в чому подібну організацію (двуслойность, променева симетрія, водний спосіб життя і т. П.), Але відрізняються за володінням НС (у губок її немає), то можна побачити, що кишковопорожнинні набагато краще адаптовані до середовища. Перш за все, це проявляється в тому, що їх тіло здатне до руху на відміну від нерухомого тіла губок. І весь подальший розвиток тварин пов'язано з дедалі більшим вдосконаленням НС. Можна уявити собі, що еволюція «спробувала» два варіанти розвитку - з НС і без неї - і «вибрала» перший.
В якості основних напрямків еволюційного розвитку НС можна виділити:
Принципово важливо, що розвиток НС в філогенезі тісно пов'язане з розвитком здатності до руху. Але швидкість рухів в процесі еволюції зростає. У безхребетних вона особливо велика у деяких членистоногих (наприклад, у двокрилих - мух, комарів і т. П.) І головоногих молюсків. Природно, що для здійснення швидких рухів необхідно підвищити швидкість поширення нервових імпульсів по рефлекторних дуг. З двох відомих для цього способів - міе- лінізаціі і збільшення діаметра нервового волокна - для безхребетних більш характерний другий з них. У ряду представників цієї групи тварин (кальмарів, ракоподібних) знайдені так звані гігантські аксони, що досягають в діаметрі майже 1 мм. У більшості випадків гігантські аксони виникають в онтогенезі тварини в результаті злиття декількох аксонів різних нейронів. Швидкість проведення в таких аксонах приблизно в 10 разів більше, ніж в звичайних аксонах того ж тварини. Цікаво відзначити, що перші класичні експерименти по реєстрації електричної активності нервових клітин були проведені саме на гігантському аксоні кальмара.
Прискорення проведення нервового імпульсу за рахунок мієлінізації також спостерігається у деяких безхребетних (комах, дощових черв'яків, крабів, креветок). Але будова мієлінової оболонки у них дещо відрізняється від уже відомих нам мієлінових оболонок хребетних. Число шарів мієліну в такій оболонці набагато менше, ніж у хребетних, вони розташовані більш пухко. Ядро гліальні клітини, що утворює оболонку, розташоване між аксонів і мієліну (рис. 16.1), а не в неврілемма, як у хребетних (див. Рис. 2.16). І хоча така оболонка, безсумнівно, збільшує швидкість проведення, але якщо порівнювати аксони однакового діаметра з мієлінових оболонками однієї товщини у хребетного і безхребетного тваринного, у першого швидкість буде значно більше.
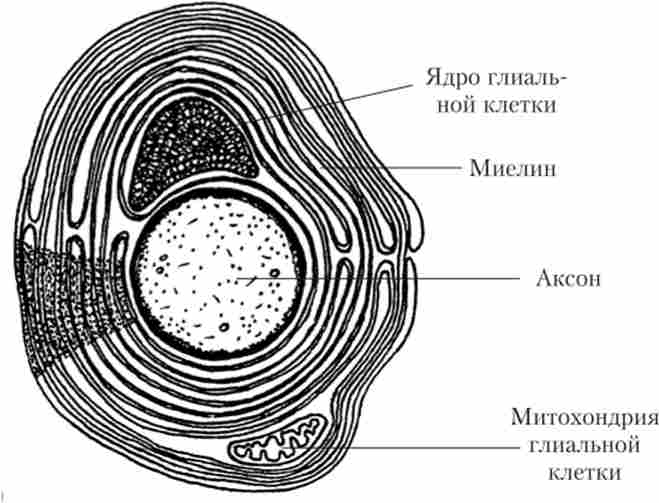
Мал. 16.1. Поперечний розріз через аксон креветки
виділяють три основні стадії в розвитку НС.
Як було відзначено, при зміні рівня розвитку на більш високий в ньому зберігаються стародавні форми організації, якщо вони можуть виконувати якусь більш приватну функцію. Так, наприклад, у хордових живіт-
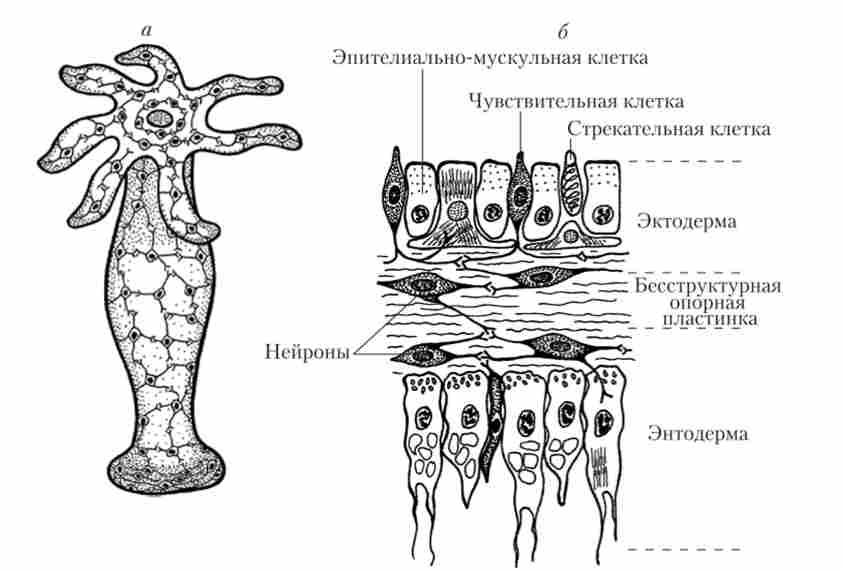
Мал. 16.2. Дифузна нервова система кишковопорожнинних:
а - НС прісноводної гідри; б - розріз через стінку тіла гідри
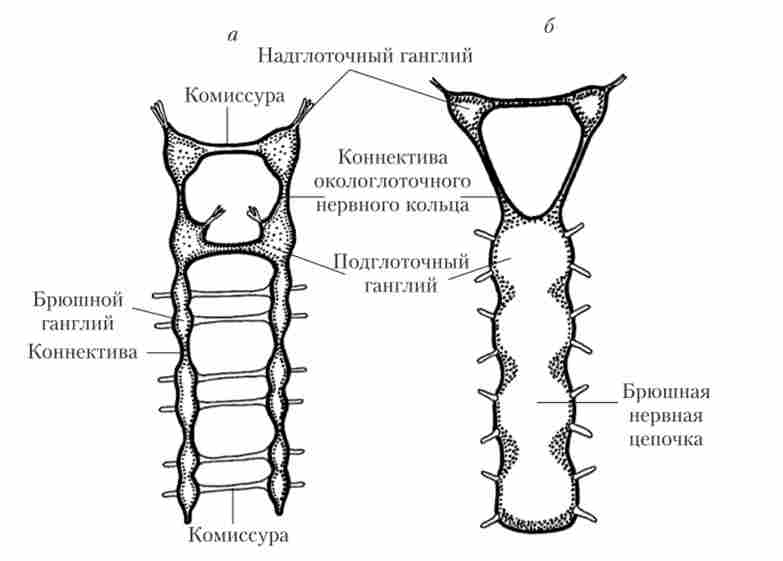
Мал. 16.3. Вузлова нервова система черв'яків:
а - сходова НС; б - НС з черевним нервовим ланцюжком
них перистальтична моторика кишкового тракту подібна моториці гідри, і тут зберігаються ділянки з дифузійної НС (інтрамуральна НС травного тракту). Інший приклад: в типі хордових спостерігається посегментно (метамерное) розташування деяких органів, більш виражене у нижчих хордових, але в якійсь мірі зберігається навіть у ссавців (хребет, СМ). І хоча ця метамерія проявляється набагато слабкіше, ніж у кільчастих хробаків і членистоногих, елементи гангліонарних НС, характерної для цих типів, у вигляді сенсорних і вегетативних гангліїв є в наявності і у хордових тварин.
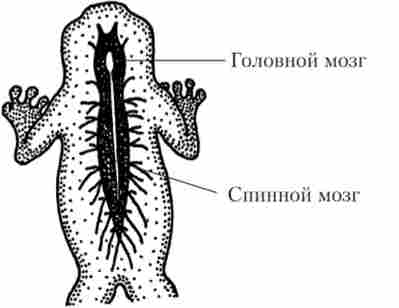
Мал. 16.4. Трубчаста нервова система жаби