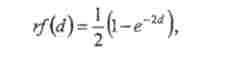
| Головна |
| «« | ЗМІСТ | »» |
|---|
Сума менших частот рекомбінації (генетичних відстаней) найчастіше перевищує частоту рекомбінації між найбільш віддаленими один від одного маркерами. Це пояснюється тим, що між будь-якими двома зчепленими генами можливий не тільки одиночний, але і подвійний (А також множинний) кроссинговеру що призводить до скорочення реєструється частоти кросинговеру. Дійсно, якби між генами b і vg не було б маркера рг, то b (pr *) vg і b+(Pr) vg * сприймалися б як некроссоверние стану b vg і b * vg+. Таким чином, подвійні обміни скорочують рег істріруемое відстань між генами.
Разом з тим між обмінами на сусідніх ділянках хромосом існує взаємовплив, назване інтерференцією. Таке взаємовплив можна виразити кількісно. Для цього зіставляють реально спостережувану частоту подвійних обмінів з частотою, теоретично очікуваної на основі припущення про те, що обміни на сусідніх ділянках відбуваються незалежно один від одного. Ступінь і характер інтерференції вимірюються величиною коінціденціі (С). Коінціденцію оцінюють як частка від ділення реально спостерігається частоти подвійних кросоверів на теоретично очікувану частоту подвійних кросоверів. Останню величину отримують, перемножая частоти кросинговеру на сусідніх ділянках.
Обчислимо коінціденцію на конкретному прикладі, користуючись даними Т. Х. Моргана і А. Стертеванта, які при трігібрідного схрещуванні вивчали рекомбінацію між генами уу w і т, локалізованими в ^ хромосомою D. melanogaster. Фенотипічніпрояв генів у і w вже описувалося. Рецесивний аллель гена т призводить до зменшення розміру крил. Частота рекомбінації між у і w становить 1,3%, а між w і т - 32,6%. Подвійні рекомбінанти по yj / vjm спостерігалися з частотою 0,045%. На підставі цих даних величина коінціденціі
С = 0,00045 / 0,013 х 0,326 = 0,00045 / 0,00424 < 1.
Величину інтерференції (/) визначають за формулою / = 1 - С.
Якщо з < 1, то інтерференція позитивна, т. е. одиночний обмін перешкоджає обміну на сусідній ділянці хромосоми. Якщо С> 1, то інтерференція негативна, т. е. один обмін як би стимулює додаткові обміни на сусідніх ділянках. Насправді існує тільки позитивна інтерференція при реципрокной рекомбінації - кроссинговере, а позірна невипадковим збіг двох і більше обмінів, характерне для дуже коротких відстаней, - результат нереці- Прокной подій при рекомбінації.
Таким чином, при картуванні генів в групах зчеплення на підставі вивчення частот рекомбінації необхідно враховувати дві протилежні тенденції. Подвійні обміни «скорочують» відстані між генами, а інтерференція перешкоджає множинним обмінам, ймовірність яких збільшується з відстанню. Як показав для дрозофіли Г. Меллер, на великих відстанях (близько 35% рекомбінації) інтерференція зникає. Отже, найбільш точні дані про частоту крос- сінговера можна отримати тільки на досить коротких відстанях - приблизно до 10 сМ.
В узагальненому вигляді залежність частоти рекомбінації від реального відстані з урахуванням множинних обмінів описує функція Дж. Холдейна:
де г / - Картира функція (у нашому випадку - частота враховуються кроссинговеров), d - реальне відстань, на якому відбуваються обміни, е - підставу натурального логарифма.
Крива, що описується функцією Холдейна (рис. 4.3), порівняна з прямою залежністю частоти реєстрованих обмінів від відстані (як це було розглянуто А. Стертевантом). Функція Холдейна показує, що зі збільшенням відстані Картира функція rf наближається до 0,5. Це означає, що між генами, розташованими далеко один від одного, виявляється близько 50 одиниць рекомбінації. Таку ж частоту рекомбінації демонструють гени, що знаходяться в різних хромосомах. Таким чином, практично неможливо вловити зчеплення між настільки віддаленими одна від одної генами. Ці гени, хоча і зчеплені фізично, перебуваючи в одній хромосомі, будуть успадковуватися незалежно.
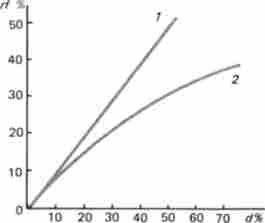
Мал. 4.3. Картира функції: 1 - лінія, описувана функцією А. Стертеванта {rf = 2 - крива, описувана функцією Дж. Холдейна (,, /? (
Доведено, що деякі гени, які контролюють 7 ознак гороху, досліджені Г. Менделем, зчеплені, проте розташовані на великій відстані один від одного. Зокрема, гени а (Забарвлення квіток і насінної шкірки) і / (забарвлення насіння) належать до однієї і тієї ж групи зчеплення, але відстань між ними - близько 200 сМ. У дослідах Менделя ці гени успадковувалися незалежно. при схрещуванні АЛІ х aaii в F2 було отримано розщеплення 357 А_1_ : 132 А Н: 116 аа1_: 34 aaii, яке добре відповідає теоретично очікуваному при незалежному успадкуванні (360: 120: 120: 40; х = 2,258; Р> 0,05).