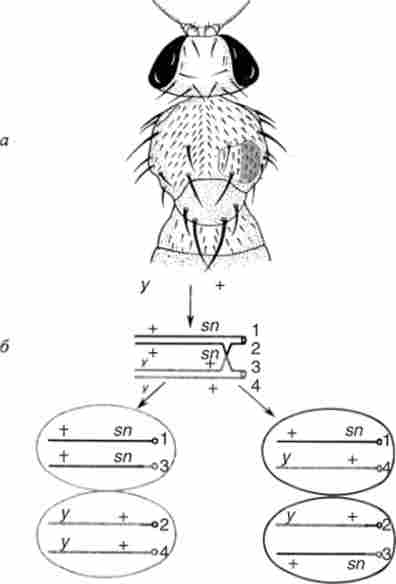
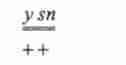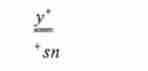Кроссинговер можливий не тільки в мейозі, а й в мітозі. У 1936 р К. Штерн досліджував мух D. melanogaster генотипу
Мутація >> - жовте тіло, sn (,singed) - обпалені щетинки. Обидва гена знаходяться в А'-хромосомі, а центромера розташована праворуч від гена sn. Досліджувані дигетерозиготи мали дикий фенотип за обома ознаками, проте зрідка на тілі деяких мух з'являлися подвійні плями: одна половина плями жовта з нормальними щетинками, інша - нормального сірого кольору, але покрита обпаленими щетинками.
Поява таких подвійних плям К. Штерн пояснив мітотичним кросинговером на стадії чотирьох хроматид на ділянці лл-центромера. Дійсно, якщо такий обмін відбудеться, то при розбіжності хромосом в мітозі в половині випадків повинні утворюватися подвійні плями (рис. 6.5).
Мал. 6.5. Митотичний кроссінговср у D. melanogaster: а - результат митотичного
у *
кросинговеру у мух, гетерозиготних по на ділянці ^ н-центромера;
* sn
б - два можливих варіанти розбіжності хромосом після мітотичного кросинговеру
?. і у *
(/, 2,3,4 - дочірні центромери); -, - утворюються в 50 % випадків
* Sn у *
Частота митотичного кросинговеру значно нижче (на 2-3 порядки) мейотичного. Проте митотичний, або соматичний, кроссинговер також можна використовувати для генетичного картування.
Припустимо, що експеримент К. Штерна було поставлено дещо інакше, і дігетерозіготние мухи нести обидва рецесивні гени в одному гомологів:
Тоді кроссинговер на ділянці. вул-центромера повинен був привести до гомозиготизації обох генів в одній клітці. Кроссинговер на ділянці y-sn повинен призводити до гомозиготизації тільки по у. оскільки ген у розташований далі від центромери, го. мозіготізація по ньому повинна відбуватися частіше, ніж по обом генам. Цей принцип використовується при генетичному картуванні у деяких грибів, у яких існує диплоидная стадія, але втрачена здатність до мейозу.
Мультиферментний комплекс - біохімія Найбільш ефективно відбувається регуляція в так званих мульти- ферментних комплексах. Ці комплекси є кілька ферментів, які каталізують ряд узгоджених реакцій, причому кінцеві продукти однієї ферментативної реакції є вихідними субстратами для наступної ферментативної реакції. Розрізняють три
Мозочок - вікова анатомія і фізіологія Мозочок розташований над довгастим мозком і мостом позаду великих півкуль головного мозку. За допомогою трьох пар ніжок - нижніх, середніх і верхніх - він з'єднується відповідно з довгастим мозком, мостом і середнім мозком. Мозочок складається з хробака і двох півкуль , поверхня яких утворює
Мозкові основи мови - фізіологія вищої нервової діяльності та сенсорних систем У 30-х рр. XX ст. У. Г. Пенфілдом була розроблена методика картування областей кори за допомогою прямого електричного роздратування. Це дозволило виявити ті структури, які найбільш важливі для здатності говорити і розуміти мову. Ця процедура здійснюється під місцевим наркозом, під час якого
Морфогенез - біологія. Частина 1 морфогенез - це процес виникнення нових структур і зміни їх форми в ході індивідуального розвитку організмів. Морфогенез, як зростання і клітинне диференціювання, відноситься до ациклічним процесам, т. е. які повертаються до свого попереднього стану і здебільшого незворотнім. Головною властивістю
Моногенне успадкування ознак. Аутосомне і зчеплене зі статтю успадкування - біологія. Частина 1 У зв'язку з тим що каріотип організму - це диплоїдний набір хромосом, більшість генів в соматичних клітинах представлені аллельними парами. Алельних гени, розташовані у відповідних ділянках гомологічних хромосом, взаємодіючи між собою, визначають розвиток того чи іншого варіанту відповідної
Молекулярні аспекти біоінженерії. Генна інженерія, загальна характеристика - біохімія частина 2. У геномі кожної живої клітини закладена генетична програма, яка визначає і контролює головні реакції клітинного метаболізму. Історія розвитку генної інженерії налічує нс більше тридцяти років. Її становлення пов'язане з конструюванням векторних молекул, отриманням рекомбінантних ДНК, а також
Множинні молекулярні форми ферментів - біохімія Участь множинних молекулярних форм ферментів в регуляції метаболізму можна проілюструвати на прикладі синтезу амінокислот у бактерій. У Е. coli аспартаткіназная реакція передує синтезу трьох амінокислот: треоніну, лізину, метіоніну. Є три ізоензіма аспартаткіна- зи (АК-1, АК-2 і АК-3), які