| Головна |
| ЗМІСТ | »» |
|---|
В результаті вивчення даного розділу студенти повинні: знати
вміти
Метаболізм. Харчування клітини. Будь-яка клітина постійно здійснює складні, різноманітні, але прекрасно скоординовані реакції перетворення речовин. Одні реакції спрямовані на виготовлення необхідних для нормальної життєдіяльності молекул {Пластичний обмін), а інші реакції, навпаки, супроводжуються розпадом харчових і допоміжних матеріалів для отримання енергії {Енергетичний обмін).
Отримувані з їжею білки, жири, вуглеводи, вітаміни і мікроелементи використовуються клітинами для синтезу необхідних їм речовин і побудови клітинних структур. Ці процеси протікають з витратою енергії. Сукупність реакцій біосинтезу речовин і їх подальшого складання в більші структури називається асиміляцією, або анаболизмом.
Переважна кількість реакцій пластичного обміну вимагає витрат енергії. Цю енергію клітини отримують за рахунок розпаду органічних речовин, які або надходять ззовні з їжею, або беруться з «запасів» клітини. При розпаді цих молекул виділяється енергія, частина якої розсіюється у вигляді тепла, а частина - запасається у вигляді молекул АТФ. При необхідності АТФ використовується для покриття енергетичних витрат клітини, в тому числі для забезпечення процесів асиміляції. Сукупність реакцій розпаду речовин, що супроводжуються виділенням і запасанием енергії, називається диссимиляцией, або катаболизмом.
Анаболизм і катаболізм - протилежні процеси: в першому випадку відбувається утворення речовин, на що витрачається енергія, а в другому - розпад речовин з виділенням і запасанием енергії. Анаболизм і катаболізм взаємопов'язані і завжди строго скоординовані і збалансовані. Порушення цього балансу призводить до розвитку захворювань або навіть загибелі, як окремих клітин, так і всього організму. Сукупність реакцій анаболізму і катаболізму, що протікають в клітині, називається метаболізмом, або обміном речовин.
Реакції метаболізму в живій клітині протікають при помірних температурах, нормальному тиску і незначних коливаннях кислотності. При таких умовах поза живих організмів ці реакції або взагалі не могли б здійснюватися, або протікали б дуже повільно. Однак в живих організмах вони протікають дуже швидко, що обумовлюється участю в них ферментів - особливих білкових каталізаторів. Так як активність ферментів дуже висока, то для підтримки нормальної швидкості метаболічних процесів потрібна незначна кількість кожного ферменту. Але ферменти діють вибірково, і клітці необхідно проводити дуже багато видів ферментів.
Всі тварини організми па Землі отримують речовини для пластичного і енергетичного обміну ззовні. Процес надходження цих речовин в клітку називається харчуванням. Одноклітинні організми самі поглинають харчові речовини, самі їх перетравлюють і самі використовують харчові молекули в різних реакціях внутрішньоклітинного обміну речовин. У багатоклітинних організмах з'їдена їжа найчастіше перетравлюється (руйнується за допомогою комплексу ферментів) в спеціально призначених для цього органах травлення, а потім з потоком крові органічні і неорганічні харчові речовини надходять до кожної клітини для засвоєння. Після перетравлення білки надходять до клітин у вигляді окремих амінокислот, вуглеводи - у вигляді моносахаридів (глюкози, фруктози), жири - у вигляді гліцерину і жирних кислот, неорганічні молекули - в розчиненому вигляді.
За рахунок енергії зруйнованих хімічних зв'язків органічних речовин спочатку синтезується універсальний для всіх живих істот джерело енергії - АТФ, який після цього витрачається на покриття енергетичних витрат.
Енергетичний обмін. АТФ синтезується в результаті реакції фосфорилювання - приєднання ще одного залишку фосфорної кислоти до молекули АДФ:
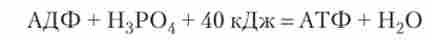
На приєднання третього фосфату до молекули АДФ витрачається 40 кДж енергії, яка зберігається в клітці у вигляді хімічного зв'язку. Такий зв'язок називається макроергічним. Якщо клітці потрібно енергія, то за рахунок розриву макроергічних зв'язку клітина отримає назад приблизно 40 кДж енергії.
Енергія для синтезу АТФ з АДФ виділяється в результаті дисиміляції, т. Е. Розщеплювання органічних речовин в клітині. Дисиміляція може проходити в два або три етапи в залежності від специфіки організму та умов його проживання (рис. 2.1).
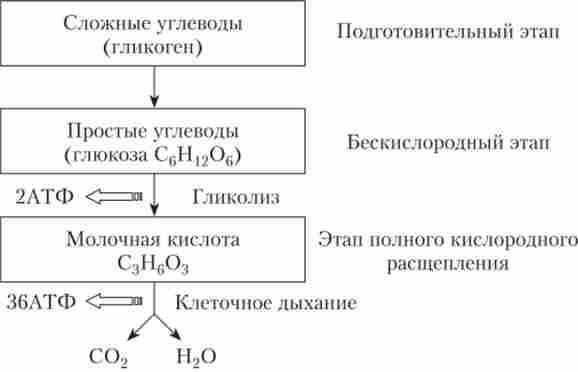
Мал. 2.1. Схема процесів енергетичного обміну
Більшість живих організмів, що мешкають на Землі, є аеробами. аероби використовують в обміні речовин кисень, що надходить з навколишнього середовища. Енергетичний обмін у аеробів протікає в три стадії: підготовча, бескислородная, киснева. У анаеробів - організмів, котрі мають потреби в кисні, а також у аеробів при дефіциті кисню енергетичний обмін відбувається в дві стадії: підготовча і бескислородная.
підготовча стадія полягає в розщепленні великих органічних молекул до більш простих: білків - до амінокислот, ліпідів - до гліцерину і жирних кислот, полісахаридів - до моносахаридів. У клітці деградація органічних речовин відбувається в фаголізосомах під дією цілого ряду лізосомальнихферментів. При цьому виділяється відносно мало енергії, що не запасається у вигляді АТФ, а розсіюється у вигляді тепла. Утворені амінокислоти, жирні кислоти і моносахариди можуть використовуватися клітиною для пластичного обміну, а також для подальшого розщеплення з метою отримання енергії.
безкиснева стадія енергетичного обміну є ферментативне розщеплення речовин, отриманих в ході підготовчої стадії. У цих реакціях кисень участі не бере. Ми розглянемо деталі другій стадії на прикладі гліколізу - безкисневого розщеплення глюкози.
гліколіз являє собою багатоступінчастий процес. Молекула глюкози, в складі якої шість атомів вуглецю (С6Н12Проб), Розщеплюється в ході гліколізу до двох молекул піровиноградної кислоти (ПВК), що містить три атома вуглецю (С3Н403). Реакції проходять в цитоплазмі клітин і катализируются комплексом ферментів. При подібному розщепленні 1 моля глюкози виділяється 200 кДж енергії, проте 60% її розсіюється у вигляді тепла. Решта 40% енергії використовується для синтезу двох молекул АТФ. ПВК в тваринних клітинах, а також в клітинах багатьох мікроорганізмів і грибів далі перетворюється в лактат - молочну киць- лоту (С3П603):
Таким чином, в результаті безкисневого ферментативного розщеплення глюкоза розпадається не до кінцевих продуктів (С02 і Н20), а до проміжних, ще багатих енергією, з'єднань. Тому в аеробних організмах після гліколізу відбувається третя, завершальна, стадія енергетичного обміну.
третя стадія енергетичного обміну є повне кисневе розщеплення, зване клітинному диханням. Органічні сполуки, що утворилися в ході другої стадії і містять запаси хімічної енергії, окислюються до С02 і Н20 - кінцевих продуктів. Цей процес також є багатостадійним, але, на відміну від гліколізу, відбувається в мітохондріях, а не в цитоплазмі. З двох молекул молочної кислоти, що утворилися на другій стадії енергетичного обміну (т. Е. З однієї молекули глюкози), в результаті повного кисневого розщеплення синтезуються 36 молекул АТФ:
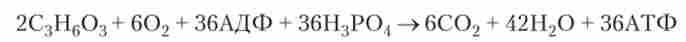
Дві молекули АТФ, крім того, утворюються в ході безкисневого розщеплення.
Таким чином, енергетичний обмін клітини в разі розпаду глюкози сумарно можна представити у вигляді такої реакції:
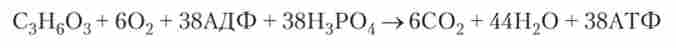
Для енергетичного обміну більшість клітин використовує вуглеводи, але для цих цілей потенційно може бути використано і окислення ліпідів, і окислення білків. Але амінокислоти, які утворюються при перетравленні білків їжі, потрібні клітці для синтезу власних білків. Тому білки і амінокислоти, що надійшли з їжею, рідко витрачаються на отримання енергії, а основним «паливом» є вуглеводи і жири.
Пластичний обмін. Синтез білків в клітині. Генетичний код. Транскрипція. Трансляція. Найважливішим елементом пластичного обміну в клітині є синтез білків. Білки виконують в клітині і організмі безліч функцій, і тому їх синтез йде безперервно. Більшість білків існує в клітці нетривалий час: в організмі людини за добу розпадається близько 0,5 кг білків, стільки ж білків синтезується заново.
Кожен вид живих істот характеризується власним, строго певним набором білків. Хоча деякі білки і виконують одну й ту ж функцію, їх набір визначає унікальність кожного виду. Білкові молекули в різних організмах можуть бути близькими за складом і навіть повністю однаковими.
Однак, всі особини одного виду хоч трохи, але відрізняються один від одного. Наприклад, всі люди схожі, але немає двох абсолютно однакових людей. Персональна неповторність забезпечується певними відмінностями в структурі білків.
Властивості білків визначаються, перш за все, послідовністю амінокислот в молекулі, т. Е. Первинною структурою білка. Інформація про первинну структуру білка кодується в нуклеотидної послідовності в молекулах ДНК. Таким чином, в ДНК міститься інформація про будову і функції як кожної окремої клітини, так і організму в цілому. Ділянка ДНК, що кодує первинну структуру одного білка, називається геном.
Кожній амінокислоті білка в молекулі ДНК відповідає послідовність з трьох один за одним розташованих нуклеотидів, які утворюють триплет, або кодон. На сьогоднішній день виявлено, які триплети в складі ДНК кодують кожну з 20 амінокислот, що входять до складу білків.
До складу ДНК можуть входити чотири азотистих підстави: аденін (А), гуанін (Г), тимін (Т) і цитозин (Ц). Число триплетів, які можуть бути складені з поєднань чотирьох азотистих основ, так само 64 (43). Таким чином, принципово ДНК могла б кодувати 64 амінокислоти. Разом з гем, встановлено, що всього кодується тільки 20 амінокислот. Пояснюється це тим, що багатьом амінокислотам відповідає не один, а кілька кодонів (табл. 2.1). Дана характеристика генетичного коду називається виродження. Вона забезпечує надійність зберігання і передачі генетичної інформації при діленні клітин. Наприклад, амінокислоті аланину відповідають чотири кодону - ЦГТ, ЦДА, ЦГГ, ЦГЦ. Випадкова заміна в третьому нуклеотиде кодону не зможе привести до змін в структурі білка - це все одно буде кодон аланина (див. Табл. 2.1).
Таблиця 2.1
генетичний код
|
перше підставу |
друга підстава |
третє підставу |
|||
|
У (Л) |
Ц (0 |
А (Т) |
Г (Ц) |
||
|
У (А) |
Фен / F Фен / F Лей / L Лей / L |
Cep / S Cep / S Cep / S Cep / S |
Тир / Y Тир / Y
|
Цис / С цис / С - стоп Три / W |
У (A) Ц (Г) A (T) Г (Ц) |
|
Ц (Г) |
Лей / L Лей / L Лей / L Лей / L |
Іро / Р Про / Р Про / P Про / P |
Гіс / Н Гіс / Н Глн / Про Глн / Q |
Apr / R Apr / R Арг / R Apr / R |
У (А) Ц (Г) А (Т) Г (Ц) |
|
А (Т) |
Ілі / 1 Ілі / 1 Ілі / 1 Мет / М старт |
Тре / Т Тре / Т Тре / Т Тре / Т |
Ach / N Ach / N Ліз / К Ліз / К |
Cep / S Cep / S Apr / R Apr / R |
У (А) U (0 А (Т) Г (Д) |
|
перше підставу |
друга підстава |
третє підставу |
|||
|
У (Л) |
Ц (Г) |
А (Т) |
Г (Ц) |
||
|
Г (Ц) |
Вал / V Вал / V Вал / V Вал / V |
Ала / А Ала / А Ала / А Ала / А |
Acn / D Acn / D Гли / Е Гли / Е |
Гли / G Гли / G Гли / G Гли / G |
У (А) Ц (Г) А (Т) Г (Ц) |
Примітка. Перший нуклеотид триплета беруть з лівого вертикального ряду, другий - з горизонтального ряду, третій - з правого вертикального;
У - урацил, А - аденін, Ц - цитозин, Г - гуанін, Т - тимін (в дужках - нуклеотид в складі ДНК, поза дужками - комплементарний йому нуклеотид в складі мРНК);
старт - старт-кодон (з нього ініціюється трансляція, т. е. починається синтез білка); стоп - стоп-кодони (на них трансляція припиняється);
амінокислоти (трибуквених російська номенклатура / однобуквеним англійська номенклатура): Феї / F - фенілаланін; Лей / L - лейцин; Ілі / 1 - ізолейцин; Мет / М - метіонін; Вал / V - валін; Cep / S - серин; Про / Р - пролін; Тре / Т - треонін; Ала / А - аланін; Тир / Y - тирозин; Гіс / Н - гістидин; Глн / Q - глутамин; АСН / N - аспарагін; Ліз / К - лізин; Acn / D - аспарагінова кислота; Гли / Е - глутамінова кислота; Цис / С - цистеїн; Три / W - триптофан; Apr / R - аргінін; Гли / G - гліцин
Ще одна властивість генетичного коду - його однозначність (специфічність). Це пов'язано з тим, що кожен триплет (кодон) визначає тільки одну амінокислоту. Так, триплет УУУ означає, що в молекулі білка буде тільки амінокислота фенілаланін. ДНК нс містить нуклеотидів, що не входять до складу будь-яких кодонів, це властивість отримало назву безперервності коду.
Молекула ДНК містить багато сотень генів, при цьому в її склад обов'язково включені особливі триплети, що позначають початок або кінець того чи іншого гена, м е виконують функцію «знаків пунктуації».
Генетичний код універсальний для всіх живих організмів, що свідчить про наявність у всіх живих істот на Землі єдиного предка.
У той час як носієм інформації про первинну структуру білків є ДНК, локалізована в клітинному ядрі, процес синтезу білка протікає в цитоплазмі, а конкретно - на рибосомах. У цитоплазму інформація про структуру білка надходить з ядра у вигляді матричної РНК (мРНК) (рис. 2.2).
При синтезі мРНК ділянку двухценочечной ДНК деспирализо, а потім на одній з ланцюжків ДНК відбувається синтез молекули мРНК за принципом комплементарності. Навпаки А молекули ДНК - У молекули РНК (замість тиміну (Т) РНК несе урацил, або У), проти Т молекули ДНК - А молекули РНК, проти Г молекули ДНК стає Ц молекули РНК і проти Ц молекули ДНК - Г молекули РНК. В результаті синтезується одна ланцюжок мРНК, яка представляє собою копію другий (нематрічной) ланцюжка ДНК, але з однією відмінністю - урацил включений замість тиміну. Завдяки такому механізму інформація про амінокислотної послідовності білка перекладається з «мови ДНК» на «мову РНК». Цей процес отримав назву транскрипції (Рис. 2.3).
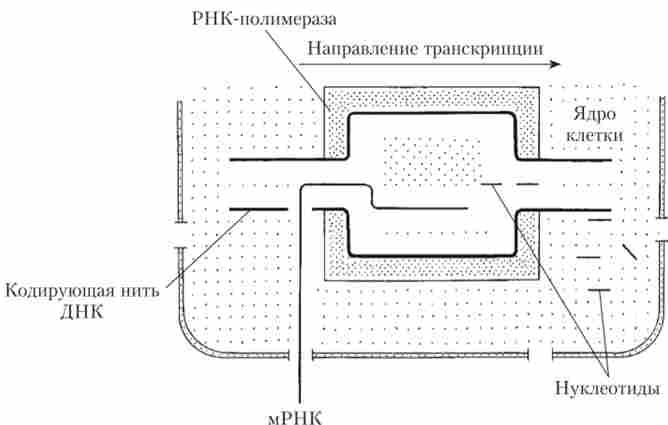
Мал. 2.2. Схема синтезу мРНК. транскрипція
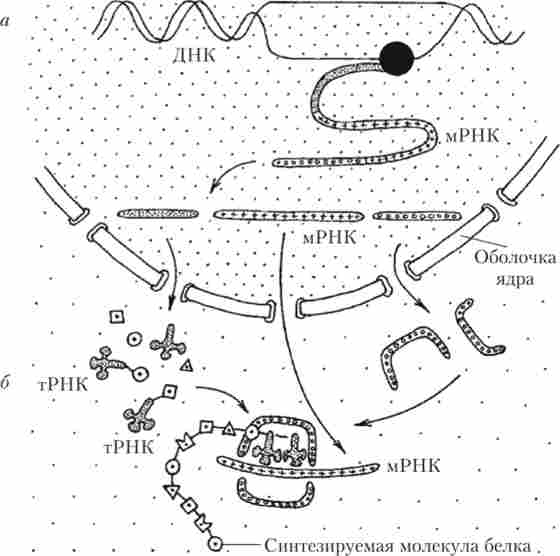
Мал. 23. Схема процесів синтезу білка:
а - транскрипція; б - трансляція
У клітині мРНК синтезується в ядрі, утворює комплекси з особливими ядерними білками і через нори в мембрані ядра переноситься в цитоплазму. У цитоплазмі повинен бути присутнім повний набір необхідних амінокислот. Велика частина амінокислот надходить в організм з травної системи, де вони з'являються в результаті розщеплювання білків, що надходять з їжею. Крім цього, деякі амінокислоти синтезуються в самому організмі. Для того щоб потрапити в рибосому і брати участь в синтезі білка, будь-яка амінокислота повинна прикріпитися до спеціальної транспортної РНК (тРНК).
«Збірка» первинному ланцюзі білка з окремих амінокислот здійснюється на рибосомах, які найчастіше розташовані на гранулярной ЕПР. Даний процес носить назву трансляції. Суть трансляції - переклад послідовності нуклеотидів молекули мРНК в послідовність амінокислот молекули білка (рис. 2.4).
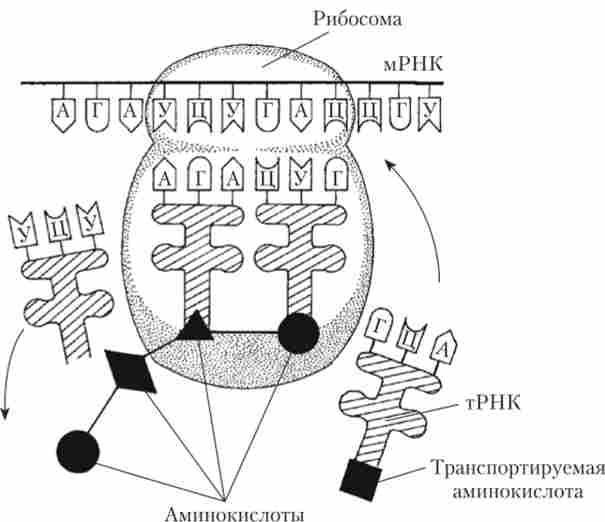
Мал. 2.4. Схема процесу трансляції:
А - адснін; Г - гуанін; У - урацил; Ц - цитозин
Рибосома взаємодіє з тим кінцем мРНК, з якого має розпочатися синтез білка, і починає рухатися по ланцюжку мРНК, затримуючись на кожному її ділянці, що включає в себе два кодони (3 + 3 = 6 нуклеотидів). Час такої затримки - 0,2 с. За цей час молекула тРНК, антикодон якої комплементарний кодону, що знаходиться в рибосомі, встигає розпізнати його. Амінокислота, яка була пов'язана з цією тРНК, відділяється від «носія» і за допомогою пептидного зв'язку приєднується до зростаючого ланцюжка білка. Одночасно до рибосоми підходить наступна тРНК, антикодон якої комплементарний наступного триплети в мРНК,
і наступна амінокислота, доставлена цієї тРНК, включається до складу зростаючої білкової ланцюжка. Рибосома після цього зсувається по мРНК, зупиняється на наступній ділянці, і весь процес повторюється заново. Нарешті, рибосома досягає одного з стоп-кодонів (УАА, УАГ або УГА) і синтез білка завершується. Всі описувані реакції йдуть дуже швидко. Підраховано, що на синтез досить великої молекули білка необхідно витратити всього близько 2 хв.
Однак клітці потрібна не одна, а багато молекул кожного різновиду білка. Тому, як тільки рибосома, першої почала синтезувати білок, просувається вперед по молекулі мРНК, на цю ж мРНК нанизується друга рибосома і починає синтез такого ж білка. Так само може бути нанизана на мРНК і третя, і четверта рибосома, і т. Д. Група рибосом, одночасно синтезують білок на одній молекулі мРНК, називають полисомой. Звільнена після закінчення синтезу білка рибосома може зв'язатися з іншою молекулою мРНК і почати синтез іншого білка, інформація про структуру якого закодована в цій молекулі мРНК. Таким чином, послідовність амінокислот в первинну структуру білка не залежить від рибосом, а визначається тільки послідовністю нуклеотидів мРНК.