| Головна |
| «« | ЗМІСТ | »» |
|---|
Перший успішний експеримент по виділенню гена, точніше, групи генів лактозного оперона (/ ас-оперону) Е. coli був виконаний в 1969 р в лабораторії Дж. Беквита. Для цього використовували два трансдуцірующіх бактеріофага X і Р80, які включили / ас-оперон в свої геноми в протилежної орієнтації по відношенню до власних генам. Оскільки підхід до виділення / ас-оперону вельми специфічний для бактерії Е. coli, його змінили інші прийоми, що застосовуються в роботі з будь-якими організмами.
Послідовності, впізнавані рестріктазамі, стохастично розкидані по геному. Чим коротше послідовність, тим частіше вона зустрічається, відповідно, тим коротше фрагменти ДНК, які утворюються при рестрикції. Ферменти рестрикції можна розділити на їхню молекулярної масі і заряду за допомогою електрофорезу в гелі. В даний час з різних мікроорганізмів виділено безліч ресгріктаз з неоднаковою специфічністю по відношенню до нуклеотидних послідовностей ДНК.
Пошук потрібного гена серед суміші рестрикційних фрагментів представляє певні складності. Поряд з цим найбільш поширеним способом отримання генів існує також спосіб хімічного синтезугенів.
Хімічний і ензиматичний синтез генів має певні переваги перед їх пошуком серед рестрикційних фрагментів. Ці методи мають і недоліки, оскільки синтетичні гени найчастіше позбавлені низки регуляторних елементів, необхідних для їх повноцінної експресії.
Клонують фрагмент ДНК може бути обмежено розмножений в ході клітинних поділів трансформанта. Вектори типу плазміди pBR322 називають векторами інактивації.
Здатність ДНК вірусу 5К40 вбудовувати в свою молекулу чужу ДНК була вивчена при отриманні гібридної молекули ДНК з двох вірусів, один з яких - онкогенний вірус SV40, а другий - дефектний фаг X з включеним в нього галактозним опероном Е. coli (Рис. 15.12). рестріктаза EcoRl розрізає кільцеві молекули вірусів в строго визначеному місці. Це переводить молекули обох вірусів з кільцевого стану в лінійне. Під дією ферменту екзонуклеаза однонітевиє ділянки розширюються на обох кінцях кожної з двох лінійних молекул. За допомогою ферменту кінцевий трансферази нарощуються кінцеві ділянки полі- (?? 4) і полі- (оТ). Тим самим створюються комплементарні однонітевиє кінці для обох молекул. Підготовлені таким чином молекули з'єднуються в кільце. Потім, застосовуючи обробку ДНК-полімеразою, забудовують однонітевиє ділянки по комплементарних матриць і за допомогою ДНК-лігази замикають гібридні кільця ковалентними зв'язками.
В результаті галактозна оперон До coli виявляється включеним в ДНК вірусу 5У40.
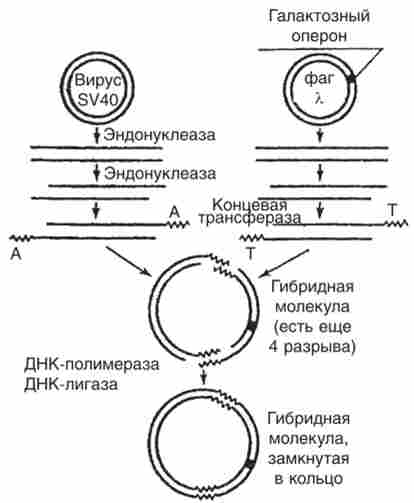
Мал. 15.12. Отримання гібридної молекули з вірусу SP40 і ДНК фага X, несучої галактозна оперон бактерії?. coli
Серед ферментів в роботах з генної інженерії головне місце займають рестріктірующіе ендонуклеази, що дозволяють отримати однорідні фрагменти ДНК, і лігази, що зшивають фрагменти. Фрагменти рестрикції кодуються хромосомними і плазмідними генами.
При розрізуванні дволанцюжкових молекул ДНК кінці фрагментів за місцем розриву можуть зберігати двухцепочечную структуру (так звані тупі кінці). ДНК-лігаза фага Т4 здатна з'єднувати тупі кінці молекул ДНК.
Для полегшення процесу зшивання на обох тупих кінцях розрізаної молекули ферментативно можна наростити липкі кінці. Такі липкі кінці, що виникають в місці розрізу двухцепочечной ДНК, здатні з'єднуватися не тільки між собою, відновлюючи вихідну кільцеву молекулу ДНК. Вони з'єднуються з будь-яким іншим фрагментом чужорідної ДНК, які мають липкі кінці, отримані при дії тієї ж рестріктази. При додаванні ферменту полінуклеотідлігази це з'єднання набуває міцний ковалентний характер. Рекомбінантна плазмида з включеним в неї ділянкою чужорідної ДНК стає тазмідним вектором. Введення в клітку і подальше розмноження рекомбінантної плазміди забезпечують клонування придбаного нею чужорідного фрагмента ДНК.
Поряд з рекстріктазамі і лігази робота по рекомбінантним молекулам вимагає цілого банку ферментів, таких як нукпеази, зворотна транскриптаза, термінальна нуклеотідінтрансфераза, ДНК-полімераза I і ін. Всі ці ферменти відонеспеціфічни, що дозволяє створювати рекомбінантні плазміди, що поєднують генний матеріал від будь-яких органічних форм.