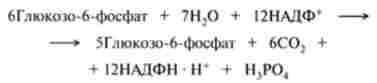
| Головна |
| «« | ЗМІСТ | »» |
|---|
Альтернативним гликолизу окислювальним шляхом катаболізму гексоз є пентозомонофосфатний, або пентозний шлях. Оскільки при цьому глюкозо-6-фосфат вимикається з метаболічного перетворення шляхом гліколізу, його також називають гексозомонофосфатного шунтом. Пентозний шлях широко поширений в природі (тварини, бактерії, рослини). В організмі людини активність цього шляху висока в клітинах лактіруюшей молочної залози, жирової тканини, зрілих еритроцитах; низький рівень цього процесу виявлено в печінці (5-10%), скелетних і серцевого м'язах (5%), мозку (10%), щитовидній залозі (15%), легких (15%).
Функції пентозомонофосфатного шляху. Цей процес виконує дві найважливіші метаболічні функції (рис. 18.8). По-перше, поставляє відновлювальні еквіваленти (НАДФН) для реакцій відновлення в процесах анаболізму, наприклад синтезу вищих жирних кислот, холестеролу і ін.
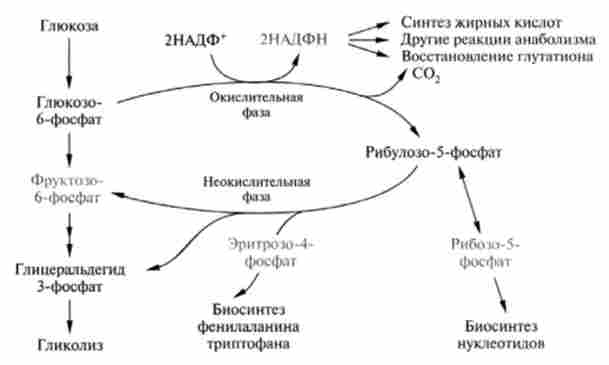
Мал. 18.8. Метаболічні функції пентозофосфатного шляху
По-друге, в пентозофосфатному шляху окислення глюкози утворюються найважливіші структурні попередники для анаболічних процесів в клітці, в тому числі рибоза-5-фосфат - для біосинтезу нуклеотидів і нуклеїнових кислот, еритроза-4-фосфат - для біосинтезу трьох амінокислот: фенілаланіну, тирозину, триптофану.
Крім цього, в аеробних умовах даний шлях може виконувати енергетичну функцію, завдяки дії ферментів, що викликають взаємоперетворенням НАДФН і НАДН. Останній, як відомо, здатний запустити процес окисного фосфорилювання для синтезу АТФ. У деяких анаеробних мікроорганізмів, наприклад облігатних гетероферментативних молочнокислих бактерій (Leuconostoc mesenteroides), Пентозний шлях є єдиним шляхом зброджування вуглеводів і, отже, забезпечення їх енергією.
У зелених рослинах метаболіти пентозного шляху виконують важливу функцію фіксації С02 в циклі фотосинтезу (гл. 16).
Реакції пентозомонофосфатного шляху. Пентозомонофосфатний шлях складається з восьми реакцій, в ньому умовно виділяють дві фази. Перша фаза - окислювальна, включає три реакції і завершується окисленням глюкозо-6 фосфату до пентозофосфатов. Друга фаза - неокислювального (П'ять реакцій), вона являє собою взаємоперетворення трьох-, чотирьох-, п'яти-, шести- і семіуглеродних сахарофосфатов, в результаті яких регенерується гексо- зо-6-фосфат. Всі реакції протікають в цитоплазмі клітини і є оборотними.
Окислювальна фаза. Вона включає наступні реакції.
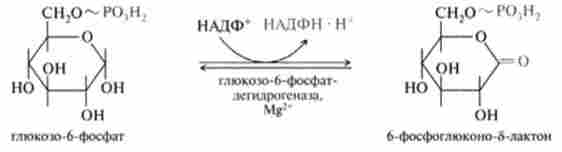
1. Реакція дегидрирования глюкозо-6-фосфату при дії глюкозо-6 фосфо! дегідрогенази, коферментом якої є НАДФ+:
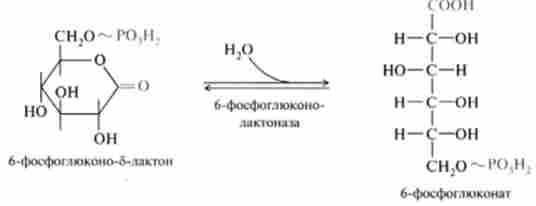
2. Реакція гідролізу лактона, яка відбувається спонтанно або за допомогою ферменту б-фосфоглюконолакгонази:
3. Реакція окисного декарбоксилювання 6-фосфоглюконат, ка
талізіруемая ферментом 6-фосфоглкжонатдегідрогеназой, НАДФ3 4-залежною, яка активується іонами Mg24. В результаті реакції утворюється перша кетопен- тоза-рибулозо-5-фосфат:
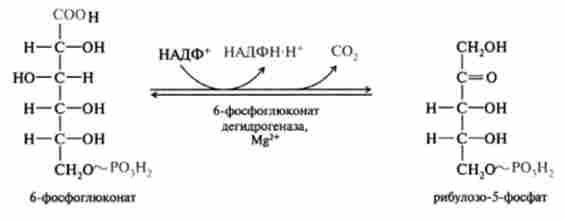
Цією реакцією завершується окислювальна фаза, в якій глюкозо-6-фосфат окислюється до рибулозо-5-фосфату і відновлюється 2НАДФН - Н+, останні використовуються як донори відновлювальних еквівалентів для анаболічних реакцій процесів метаболізму. Стехіометричне рівняння окисної фази пентозофосфатного шляху описується рівнянням
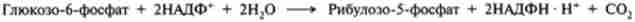
Неокислювального фаза. Основними ферментами неокислювального фази є два ферменти: транскетолаза і трансальдолаза. Вони отщепляют від фосфорілірованний кетосахаров відповідно З2- і С3-фрагменти, переносячи їх на фосфоальдосахара. В результаті взаємодії фосфосахаров регулюється їх кількість відповідно до потреб клітини.
Неокислювального стадія починається з реакцій ізомеризації (4) і (5). В процесі цих реакцій одна частина рибулозо-5-фосфату изомеризуется в ри- бозо-5-фосфат (акцептор З2-фрагмснта), інша - в ксилулозо-5-фосфат, який служить донором цього фрагмента.
Якщо рибоза-5-фосфат втягується в анаболічні процеси, пентозний шлях на цьому етапі може завершуватися.
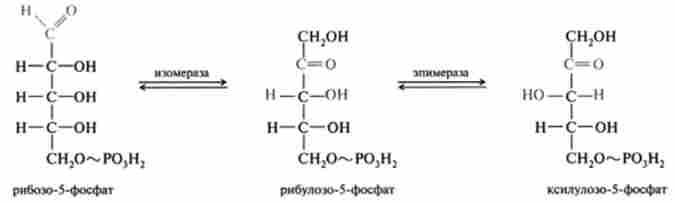
6. Перша транскетолазная реакція йде між продуктами четвертої і п'ятої реакцій при дії ферменту транс кетолази, кофакторами якого є тіамін пірофосфат (ТПФ) і катіони двовалентних металів. Реакцію можна розглядати як оборотний перенесення глікоальдегідного залишку від донора кетози на акцептор альдозами:
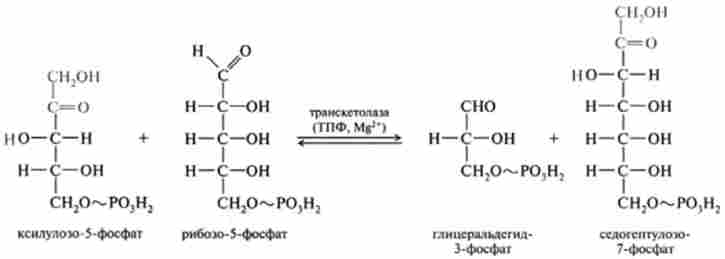
7. Трансальдолазная реакція протікає за допомогою ферменту трансаль- долази, який каталізує перенесення залишку діоксіацетон від донора кетози, представленої седогептулозо-7-фосфатом, на акцептор альдозами - гліце- ральдегід-3-фосфат:
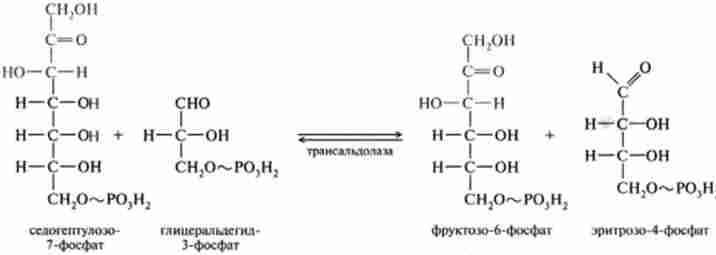
8. Друга транскетолазная реакція, в якій донором глікольальдегід- ної групи виступає, як і в шостий реакції, ксилулозо-5-фосфат, акцептором - продукт сьомий реакції еритроза-4-фосфат:
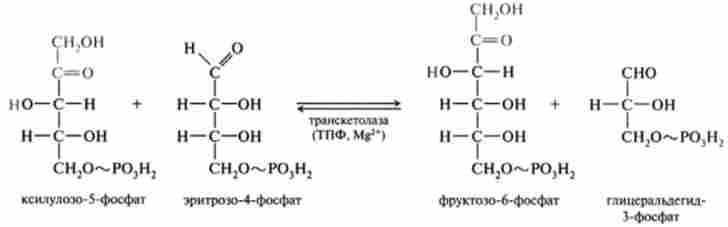
Таким чином, перша транскетолазная реакція між двома фосфопен- тозамі відкриває серію реакцій перетворення моносахаров (6), (7), (8), які можна представити у вигляді такої схеми:
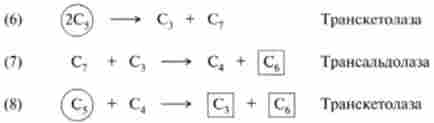
В результаті цих реакцій три молекули С5-цукрів (обведені в кружок) перетворюються в дві молекули С()-цукрів і одне З3-з'єднання - гліцеральде- гід-3-фосфат (обведені в квадрат). Цей процес можна описати рівнянням
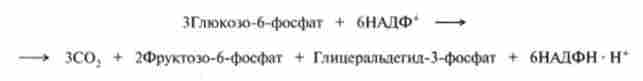
Всі кінцеві продукти цих перетворень можуть далі метаболизировать по шляху гліколізу, забезпечуючи як енергетичні потреби клітини, так і потреби в НАДФН (рис. 18.9).
Якщо ж клітина не потребує НАДФН. а їй потрібен рибоза-5-фосфат для синтезу нуклеотидів, то проміжні метаболіти гліколізу - фрукто- зо-6-фосфат і 3-фосфогліцеріновий альдегід - при дії ферментів неокислювального фази пентозофосфатного шляху можуть перетворюватися в рибоза-5-фосфат. Схема цього перетворення наведена на рис. 18.10.
Таким чином, пентозофосфатний шлях відрізняється крайньою гнучкістю. Якщо в клітці потреби в рибоза-5-фосфат і НАДФН збалансовані, то неокислювального стадії не потрібно. Але якщо, наприклад, жирових клітин НАДФН потрібно значно більше, ніж рибоза-5-фосфату, то надлишок останнього в результаті реакцій неокислювального фази перетворюється в глюко- зо-6-фосфат:
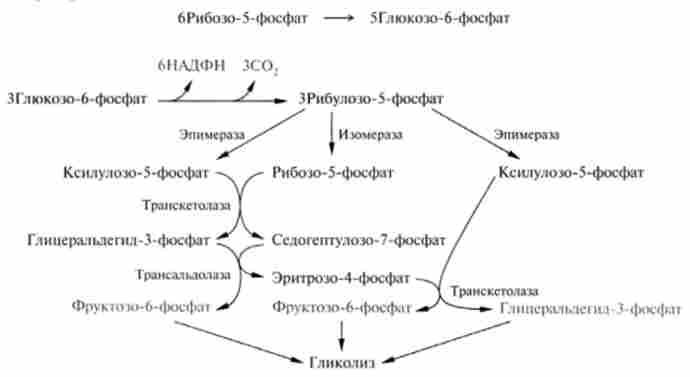
Мал. 18.9. Схема реакцій пентозофосфатного шляху
За допомогою існтозофосфатного шляху може відбуватися повне окислення глюкозо-6-фосфату до С02, якщо провести розрахунок на шість молекул глюкозо-6-фосфату, які утворюють п'ять молекул глюкозо-6-фосфату і шість молекул С02 за рівнянням
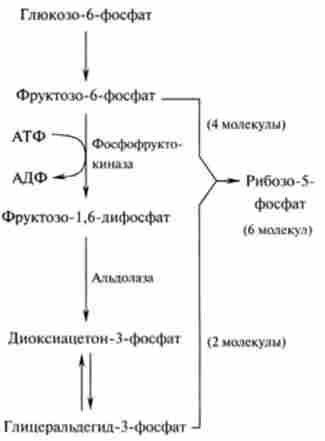
Мал. 18.10. Схема перетворення метаболітів гліколізу в рибоза-5-фосфат
Після скорочення загальних членів отримуємо:
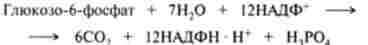
Однак слід пам'ятати, що всі шість молекул С02 утворюються з С-1-атомів шести молекул глюкозо-6-фосфату, а з утворених при цьому шести молекул рибулозо-5-фосфату знову регенеруються п'ять молекул глюкозо-6-фосфату.
Регуляція пентозомонофосфатного шляху. Головним регуляторним ферментом пентозного шляху є глюкозо-6-фосфатдегідрогенази, що каталізує першу реакцію. Активатором ферменту є НАДФ а роль негативного ефектора (інгібітора) виконує відновлена форма цього коферменту НАДФН - Н Чутливий до співвідношення | НАДФ *) / [НАДФН - Н+1 і другий окислювальний фермент - 6-фосфоглюконатдегід- рогенеза. При величині співвідношення 0,02 їх активність максимальна, при 0,01 вона зменшується на 90%.
У перемиканні пентозного шляху і гліколізу один на одного роль регулятора виконує еритроза-4-фосфат. Якщо пентозофосфатов багато, то еритро- зо-4-фосфат бере участь в транскетолазной реакції, що призводить до утворення фруктозо-6-фосфату і його альдоізомера глюкозо-6-фосфату. Якщо ж багато гексозофосфат, то еритроза-4-фосфат вступає в трансальдолазную реакцію, поповнюють пул седогептулозо-7-фосфату.