| Головна |
| «« | ЗМІСТ | »» |
|---|
Один з небагатьох (якщо не єдиний) прикладів детермінації стосується визначення клітинного типу (типу спарювання) у дріжджів Лгсс / г. сегеутае. Це доводить можливість перебудови генетичного матеріалу в онтогенезі.
Як уже згадувалося, у гомоталлічних дріжджів гаплоїдні аско- суперечки - продукти мейозу - дають початок диплоїдним культурам. Це здійснюється завдяки переключенню типів спарювання а<=^ а з імовірністю, близькою до одиниці, при перших діленнях проростає аскоспори. Перемикання відбувається згідно з так званим касетного механізму, запропонованого в кінці 1970-х рр. А. Херсковіц, Дж. Хіксом і Дж. Стразерном. Поряд з локусом типів спарювання (МАТ) поблизу центромери III хромосоми у дріжджів є дві мовчазні «Касети», містять неекспрессіруемие аллели в лівому плечі (НМЬа - для (а-типу спарювання) і в правому плечі (НМ! ^, - для а-типу спарювання) тієї ж хромосоми.
При перших діленнях аскоспори тип спаровування перемикається на протилежний під контролем гена АЛЕ (Від англ. Іото-Лаш. чт). ген АЛЕ кодує ендонуклеази, яка виробляє двунітевой сайт- специфічний розріз ДНК в локусі МАТа, або МАТ "в залежності від того, який аллель присутній в цьому локусі. двунітевой розріз ініціює спрямовану конверсію, при якій генетична інформація касети НМТа заміщає інформацію, що міститься в локусі МАТЙ (Або НМДз заміщає інформацію МАТа). При цьому касети зберігають міститься в них генетичний матеріал, а генетичний матеріал, який перебував в локусі МАТ, втрачається. Таке перемикання відбувається тільки в двох клітках на стадії мікроколонії, що складається з чотирьох клітин. Після цього клітини типу спарювання а копуліруют з клітинами типу спарювання а. Утворюються диплоїдні клітини, гетерозиготні по МАТй/ МАТа, і ген АЛЕ вимикається. Далі гетерозиготний диплоид стабільно розмножується до нового мейозу і споруляции, після чого при ізоляції аскоспор весь процес в ході їх проростання повторюється.
Як показали експерименти з клонування і гібридизації ДНК ДНК, локус МАТ і касети НМЬа і НМКа містять як однакові, так і різняться нуклеотидні послідовності. як МАТ0, так і МАТа утворюють по два протилежно спрямованих транскрипта, при цьому встановлено, що принаймні в разі МАТа транскрипція правого (а1) і лівого (А2) ділянок відбувається з загального промотора розміром в 10 п. н., розташованого між ними (рис. 10.17):
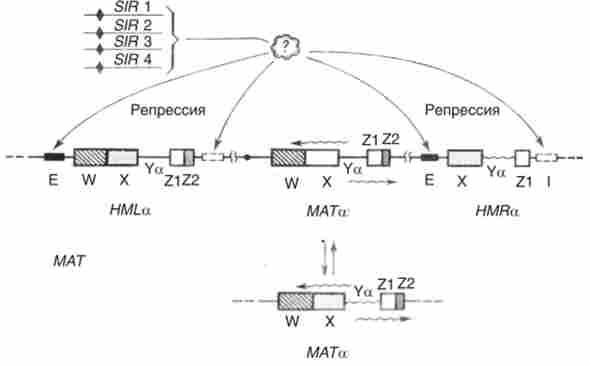
Мал. 10.17. Структура касет і локусу МАТ у дріжджів Saveli, cerevisiae. Однакові послідовності (.X, Y, Z) дані однаковим кольором; Еп1- глушники; хвилястими лініями - транскрипти; гени SIR контролюють репресію касет
Інформація, укладена в касетах, які не експресується внаслідок того, що кожна з них фланкирована послідовностями - «глушниками», що впливають на характер компактизации хроматину в касетах. Глушники функціонують під контролем кількох генів, названих SIR (silent information regulators - регулятори мовчить інформації), локалізованих в інших хромосомах. Близько локусу МАТ глушників немає.
Власне, перемикання інформації локусу типу спарювання, т. Е. Переміщення касети, і являє собою акт детермінації клітинного типу. Подальша диференціювання типу спарювання полягає в специфічній регуляції цілих серій генів: ^ -специфічні або а-специфічних. На думку авторів касетної моделі, подібні багатоступінчасті взаємодії регуляторних і структурних генів можуть забезпечувати досить складні акти детермінації у багатоклітинних організмів.
Подібний механізм касетної регуляції типів спарювання знайдений в іншого роду дріжджів - Schiz. potnbe. Касетний механізм бере участь в перемиканні поверхневих антигенів трипаносом і гонококів. Життєвий цикл гомоталлічних сахароміцетів дуже схожий з життєвим циклом мохів та лишайників. Таким чином, касетний механізм регуляції онтогенетичних змін, очевидно, набув значного поширення в природі.