| Головна |
| «« | ЗМІСТ |
|---|
Прикладом перебудови генетичного матеріалу в онтогенезі є диференціювання лімфоцитів при розвитку системи імунітету у хребетних тварин і людини.
антитіла, виробляються імунною системою, пов'язують потрапляють в організм чужорідні білки, полісахариди і ряд інших з'єднань, які називаються антигенами. Ссавці можуть продукувати до 106 різних антитіл. «Розпізнавання» антигенів забезпечують два основні класи лімфоцитів - клітин, що утворюються в кістковому мозку: Т-лімфоцити, локалізовані в зобній залозі (тимусі), і В-лімфоцити, диференціюються в селезінці і лімфоїдних органах.
Біосинтез антитіл здійснюють В-лімфоцити, які взаємодіють з Т-лімфоцитами. Антиген зв'язується з рецепторами на поверхні як В-, так і Т-лімфоцитів, після чого Т-лімфоцит «дозволяє» В-клітинам синтез імуноглобулінів, т. Е. Антитіл. Лімфоцити реагують на величезне число антигенів і «запам'ятовують» кожен з них так, що при повторному їх попаданні в організм утворюються імуноглобуліни пов'язують саме цей антиген. Генетичний контроль імунної відповіді вивчає область генетики, названа імуногенетика.
Кожна молекула імуноглобуліну - це димер, що складається з двох важких (Н) і двох легких (?) ланцюгів (рис. 10.18). Відомо злоякісне утворення імунної системи - миелома, при якій відбувається розмноження клітин, що продукують антитіла тільки одного типу. Порівняння імуноглобулінів з різних мієлом мишей і людини показало, що як # -, так і? -Ланцюга мають константні і варіабельні ділянки (рис. 10.18). Центр зв'язування антигену утворюють варіабельні ділянки Н і? -ланцюгів.
Загадка співіснування константних і варіабельних ділянок в одній і тій же поліпептидного ланцюга була дозволена в кінці 1970-х рр. С. Тонегава. Фрагменти ДНК, що кодують V- і С-ділянки, розташовані у вигляді безперервних послідовностей в геномі мишачої мієломи, просторово розділені у ембріонів або в сперматозоїдах миші. В ході диференціювання лімфоцитів гени, що кодують V- і С-ділянки, перебудовуються таким чином, що в результаті виявляються частинами одного і того ж гена, транскрібіруемих як ціле.
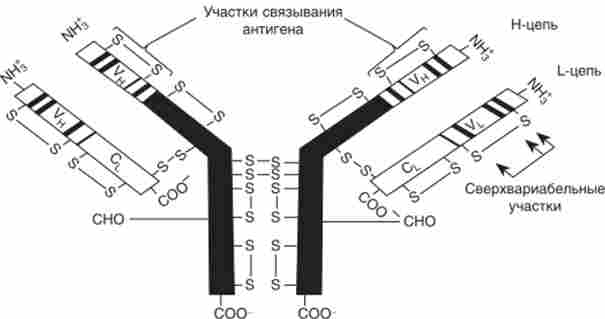
Мал. 10.18. Схема будови молекули людського імуноглобуліну; У і С- варіабельні і константні ділянки поліпептидних ланцюгів; Н і? - важкі і легкі ланцюги
Доказ перебудови генів, що кодують V- і С-ділянки імуноглобулінів, отримано завдяки техніці рекомбінантної ДНК. Інформаційну РНК для легкого ланцюга виділили з полісом мієломи і отримали кДНК за допомогою зворотної транскриптази. Препаративне кількість цієї кДНК було «напрацьовано» шляхом клонування в складі вектора бактеріальних клітин. Очищену від вектора кДНК, що кодує? -Ланцюг імуноглобуліну, розділили рестриктазой на фрагменти, відповідні К і С-ділянкам. Препарати ДНК позначили радіоактивним фосфором і використовували в якості зондів для гібридизації з ДНК мієломи і ембріона миші (рис. 10.19).
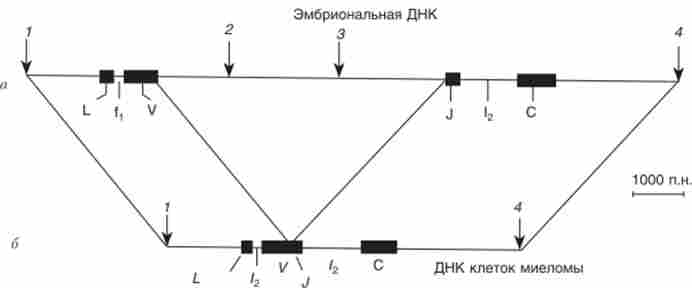
Мал. 10.19. Розташування ділянок ДНК, що кодують?-Легені ланцюги імуноглобулінів: а - в ембріоні миші; б-в - миеломе миші: 1-4 - сайти рестрикції ЕсоЯI; іРНК легкого ланцюга містять лідер (?), ділянка, що кодує вариабельную ланцюг (У і 7), і ділянку, що кодує константну ланцюг (С); ДНК мієломи зберігає тільки сайти рестрикції 1 і 4 і включає два інтрони 1 і /2
Виявилося, що в одному фрагменті, отриманому з ДНК мієломи ре- стріктазой ЕсоШ, містяться послідовності, що кодують як вариабельную, так і константну частини? -ланцюга (рис. 10.19). У той же час в ембріональної ДНК вони входять до складу різних? зі /? 1-фрагментів. Отже, в онтогенезі генетичний матеріал перебудовувався, що призвело до зникнення рестрикционного сайту ЕсоШ і зближенню раніше розділених ділянок ДНК.