| Головна |
| «« | ЗМІСТ | »» |
|---|
Азот амінокислот, відщеплюється, як правило, на ранніх стадіях катаболізму, включається до загального метаболічний пул. Залежно від потреб організму він може рсутілізіроваться в анаболічних процесах або включатися в кінцевий продукт обміну азоту - сечовину і виділятись з організму.
Безазотисті вуглецеві залишки амінокислот утворюють кислоти, найчастіше за все кетокислот, які далі деградують за загальними шляхах катаболізму інших окислених вуглеводнів (рис. 24.7).
Метаболіти, які утворюються з вуглецевих скелетів амінокислот, або безпосередньо включаються в цикл трикарбонових кислот, або перетворюються в піруват і через ацетил-КоА деградують до утворення кінцевих продуктів - С02 і Н20. У залежності від потреб організму безазотистих метаболіти можуть включатися в синтез глюкози (Гликогенние амінокислоти) або в синтез вищих жирних кислот (Кетогенні амінокислоти).
До глікогену амінокислот відносяться тс амінокислоти, при катаболизме яких утворюються безпосередні попередники глюкози, залучаємо до процесу глюконеогенезу - піруват, оксалоацетат, фосфоеноілпіруват (таких амінокислот 14), або в жири (кетогенні, одна амінокислота), або і в вуглеводи, і в жири ( гликогенние і кетогенні, 5 амінокислот). Таким чином, класифікація
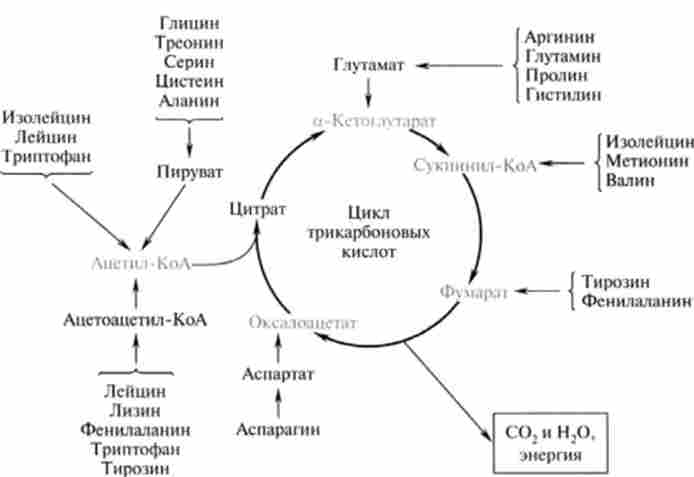
Мал. 24.7. Метаболіти, які утворюються з вуглецевих скелетів амінокислот
амінокислот за ознакою кстогснності або глікогену досить умовна, оскільки інтермедіати деяких з них можуть включатися як в синтез вуглеводів, так і жирів (табл. 24.2).
На рис. 24.8 представлені шляхи окисного розпаду амінокислот з розгалуженим ланцюгом - кетогенной амінокислоти лейцину, а також валина і ізолейцину, які є одночасно кетогенная і глікогену. В процесі метаболічних перетворень валина відбувається утворення сукцинил-КоА, який через цикл ТКК і за участю деяких інших ферментів може перетворитися в піруват, а потім в глюкозу. У той же час лейцин дає безпосередньо кетопродукт ацетоацетат і, крім того, аце- тил-КоА, з якого також може утворюватися ацетоацетат. Ізолейцин дає аіетіл-КоА і пропіоніл-КоА. Через метил малої іл-КоА пропи- ОНИЛ-КоА перетворюється в сукцинил-КоА, і, отже, його слід вважати глікогену, а так як ацетил-КоА - кетогенная з'єднання, то изолейцин можна віднести одночасно до обох категорій.
Таблиця 24.2. Гликогенние і кетогенні амінокислоти
|
гликогенние |
кетогенні |
Гликогенние і кетогенні |
|
|
аланин |
гістидин |
лейцин |
ізолейцин |
|
аргінін |
метіонін |
лізин |
|
|
аспарагінова кислота |
пролин |
феніл аланин |
|
|
цистеїн |
Серін |
тирозин |
|
|
глутамінова кислота |
треонін |
триптофан |
|
|
Глінін |
глутамин |
||
|
аспарагин |
валін |
||
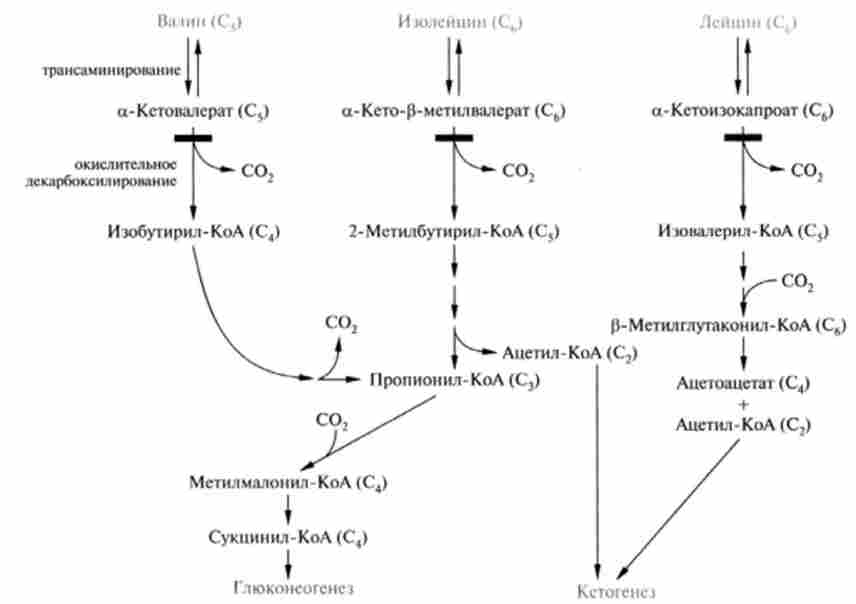
Мал. 24.8. Шляхи катаболізму амінокислоті розгалуженим ланцюгом
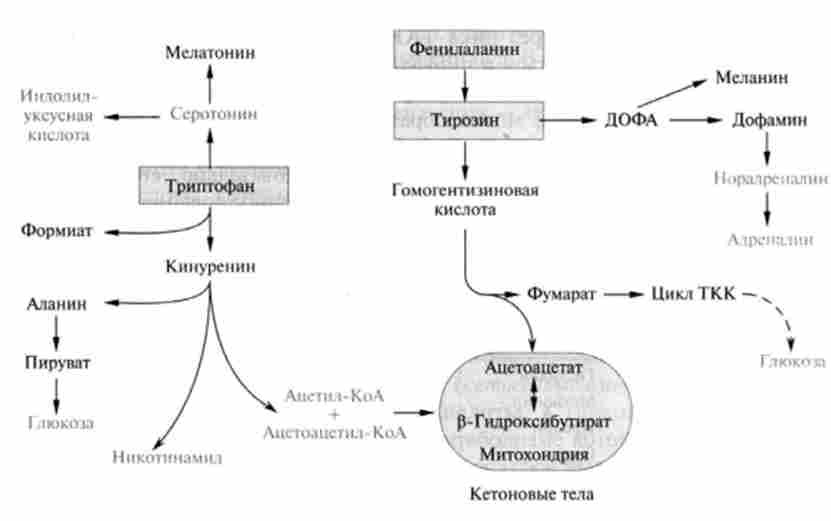
Мал. 24.9. Шляхи катаболізму ароматичних амінокислот
До групи амінокислот, безазотистих залишки яких здатні метабо- лизировать як по шляху вуглеводного, так і ліпідного обміну, відносяться ароматичні амінокислоти - триптофан, фенілаланін, тирозин, схема шляхів катаболізму яких приведена на рис. 24.9.
Метаболічні перетворення триптофану призводять до утворення різних важливих дзвінків, в тому числі серотоніну - речовини, присутнього в нервовій, а також в деяких інших тканинах ссавців, що володіє потужним вазопресорних дією; индолилуксусной кислоти - ростового речовини рослин (типу ауксину); вітаміну нікотинової кислоти, необхідної для синтезу ні коти нам ід них коферментів (НАД * і НАДФ+).
Катаболізм триптофану в організмі йде в основному по кінуреніновому шляху (близько 95%) і лише незначною частиною - по серотон і новому, в процесі якого утворюється індолілуксусная кислота, яка виводиться з організму з сечею.
Основні метаболіти, що утворюються в процесі катаболічних перетворень природних амінокислот, наведені нижче:
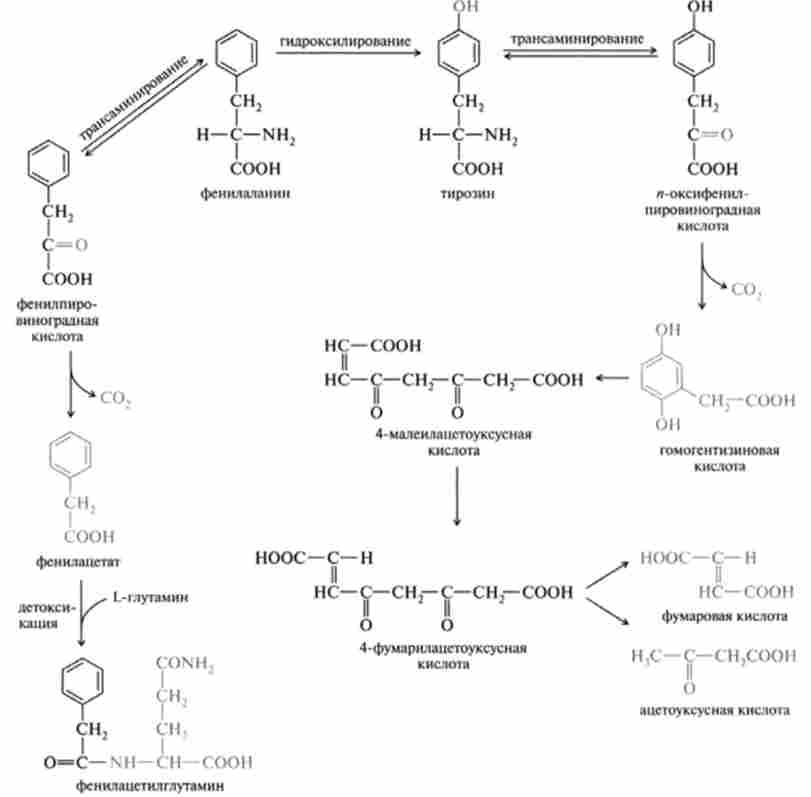
Слід зазначити, що окислювальний розпад природних амінокислот представляє особливий інтерес у зв'язку з тим, що багато вроджені порушення білкового обміну пов'язані саме з обміном цих амінокислот, наприклад спадкова хвороба фенілкетонурія (Фенилпировиноградная олігофренія). Причиною цього захворювання є втрата здатності організму синтезувати фермент фенілаланін-4-монооксігеназу. каталізують перетворення фенілаланіну в тирозин. Це призводить до накопичення фенілаланіну в тканинах, а отже, і продуктів його трансаминирования фенілпіро- виноградної і фенилуксусной кислот, що надають токсичну дію на організм, і в першу чергу на центральну нервову систему, викликаючи розлад психічної діяльності людини.
Розпад тирозину починається з трансаминирования, що приводить до утворення л-оксіфенілпіровіноградной кислоти. Остання при дії ферменту оксидази л-оксіфенілпіровіноградной кислоти (медьсодержащий білок) каталізує перетворення свого субстрату в гомогентизинової кислоту, причому реакція ця включає в себе декарбоксилирование, окислення, гідроксилювання ароматичного ядра і міграцію бічного ланцюга. Оксілаза гомогентизиновой кислоти - фермент, який потребує іони двовалентного заліза, каталізує утворення з гомогентизиновой кислоти малеілаіе- тоуксусной кислоти. Остання перетворюється в відповідне транссосді- ня - фумарілацетоуксусную кислоту, розщеплення якої призводить до утворення, по-перше, фумарової кислоти, яка піддається подальшим перетворенням за участю ферментів циклу ТКК, і, по-друге, ацето- оцтової кислоти, яка може або увійти також в цикл ТКК (у формі ацетил-КоА), або перетворитися в жирні кислоти.