| Головна |
| «« | ЗМІСТ | »» |
|---|
Основними властивостями серцевого м'яза, що визначають безперервне ритмічне скорочення серця протягом усього життя організму, є автоматия, збудливість, провідність і скоротність.
Автоматия. Під автоматией розуміють здатність серцевого м'яза ритмічно порушуватися і скорочуватися без будь-яких зовнішніх по відношенню до серця впливів, т. Е. Без участі нервової системи і гуморальних факторів, що доставляються до серця кров'ю.
Доказом автоматии серця послужили наступні спостереження та експерименти.
Ізольоване серце, т. Е. Виведене з організму і поміщене в живильний розчин, продовжує мимовільно скорочуватися. Навіть розрізане на шматочки, воно скорочується в тому ж ритмі, що й у здорової тварини. Якщо у тварини денер- вати серце, т. Е. Перерізати всі нервові стовбури, які підходять до серця, воно продовжує скорочуватися.
На здатності працювати без впливу зовнішніх подразників заснована пересадка серця. Пожвавлення зупиненого серця досягається відновленням спонтанної активності серця, його автоматии.
У чому причина такого унікального властивості серця? У більшості безхребетних тварин автоматия пов'язана з нервовими гангліями, розташованими поблизу серця, т. Е. Має нейрогенную природу. У всіх же хребетних тварин і у частини безхребетних автоматия серця обумовлена не нервовими, а м'язовими клітинами, які мимоволі деполярізуется після кожного потенціалу дії. Ці клітини називаються Пейсмекер, або «задають серцевий ритм», або водіями серцевого ритму. Така теорія автоматии серця називається миогенной.
Здатністю до автоматии мають атипові м'язові клітини, складові провідну систему серця.
Провідну роль в автоматии грає синусний вузол. Він має найвищу активність в порівнянні з іншими ділянками провідної системи, частота імпульсації в ньому найбільш висока, і він задає певну частоту скорочення серця в стані фізіологічного спокою. Такий ритм зазвичай називають Синусно ритмом, а синусний вузол - водієм ритму серця першого порядку.
Якщо відокремити лигатурой синусний вузол від передсердь (досвід Станниуса), то зазвичай серце зупиняється. Однак через деякий час воно знову починає скорочуватися, але в більш рідкісному ритмі. Цей ритм «задає» наступний вузол провідної системи-атріовентрикулярний. Більш рідкісні скорочення серця обумовлені тим, що збудливість атріовентрикулярного вузла менше, ніж синусового. Цей вузол називають водієм ритму серця другого порядку. Якщо ж і атріовентрикулярний вузол перестає генерувати збудження, то водієм ритму серця стає пучок Гіса, але його збудливість ще менше; пучок Гіса називають водієм ритму третього порядку.
У звичайних умовах атріовентрикулярний вузол і пучок Гіса тільки проводять збудження від синусового вузла. Їх власна автоматия як би пригнічена головним пейсмекером, і тільки при розвитку патологічного процесу, який припиняє функцію синусового вузла, свій ритм нав'язують нижележащие вузли. Вони є латентними, або прихованими, або потенційними Пейсмекер.
Яка природа автоматии? Методами електрофізіології встановлено, що потенціал дії (ПД) клітин провідної системи відрізняється від інших м'язових і нервових клітин. Під час розслаблення серця - діастоли - починається повільно наростаюча деполяризація мембрани, яка потім переходить в фазу швидкого деполяризації (рис. 6.3, А). Фаза реполяризації в пейсмекером досить тривала, в пейсмекером синусового вузла вона має виражену плато замість піку потенціалу. Відразу після повернення мембранного потенціалу до рівня потенціалу спокою знову починається повільна діастолічна деполяризація мембрани, і коли різниця потенціалів між зовнішньою і внутрішньою поверхнями мембрани зменшується до певного критичного, або порогового рівня, раптово виникає новий крутий зрушення електричного заряду клітини, що свідчить про її порушення.
Інтервал між двома ПД залежить від тривалості повільної діастолічної деполяризації, її величини і порогового рівня серцевого ПД. Якщо швидкість деполяризації зменшує-
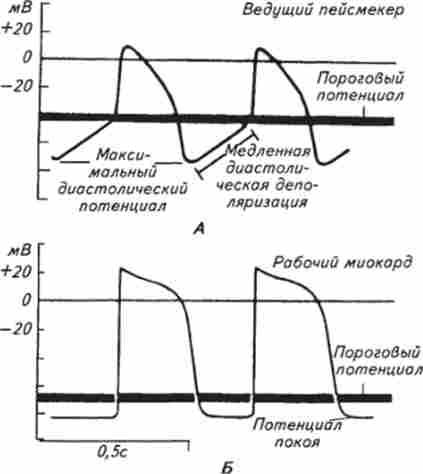
Мал. 6.3. Електричний потенціал пейсмекера {А) і робочого міокарда (Б)
ся (наприклад, при охолодженні синусового вузла), то граничний рівень деполяризації настає пізніше, частота ПД і скорочень серця зменшуються. При зростанні швидкості деполяризаціїмембрани, навпаки, пороговий рівень деполяризації виникає раніше і це призводить до почастішання порушення серця. Почасти цим пояснюється почастішання серцевої діяльності при підвищенні температури тіла.
Повільна діастолічна деполяризація обумовлена особливостями іонної проникності мембрани пейсмекеров. Як і в інших клітинах, електричні процеси в мембранах міокарда є наслідком пасивного і активного переміщення іонів натрію і калію через найтонші канали (пори) в мембрані, проникність яких регулюється зарядженими частинками - іонами Са2+ або Мп2 . Повільна діастолічна деполяризація пояснюється тим, що під час реполяризації частина натрієвих каналів не інактивується і здійснюється повільний вхід спочатку натрію, а потім кальцію в мембрану. Коли кількість іонів натрію, що проникли в клітку, знизить мембранний потенціал до критичного рівня, настає швидка фаза деполяризації і ПД досягає свого максимального рівня.
В теорії про автоматии пейсмекеров ще багато неясного, і розкриття найтонших механізмів електричних процесів, що відбуваються в серці, - актуальне завдання сучасної кардіології.
Збудливість. Збудливість - властивість серцевого м'яза переходити в стан збудження під впливом різних подразників.
У природних умовах подразником є ПД, що виникає в синусному вузлі і поширюється по провідній системі серця до робочих кардіоміоцитів. При деяких захворюваннях серця роздратування може виникати в інших його ділянках, які генерують власні ПД, і тоді серцевий ритм буде порушений через взаємодію різних за частотою і фазою ПД. В експериментах на тваринах як подразників можуть бути використані механічні, термічні або хімічні впливи, якщо їх величина перевищує поріг збудливості серця.
При хворобах серця, що супроводжуються порушенням серцевого ритму, хворим імплантують в серці мініатюрні електроди, що живляться від батарейок. Імпульси струму подаються безпосередньо на серце і збуджують в ньому ритмічні імпульси. При раптовій зупинці серця або порушення синхронізації окремих м'язових волокон можливий вплив на серце прямо через шкірний покрив сильним коротким електричним розрядом напругою в кілька кВт. Це викликає одночасне збудження всіх м'язових волокон, після чого відновлюється робота серця.
Під час збудження в серці виникають фізико-хімічні, морфологічні та біохімічні зміни, які призводять до скорочення робочого міокарда. Одними з ранніх ознак порушення є активація натрієвих каналів і дифузія іонів натрію з міжклітинної рідини через мембрану, що призводить до її деполяризації і виникнення ПД.
У клітинах робочого міокарда ПД дорівнює 80 ... 90 мВ, при ПД 100 ... 120 мВ повільна діастолічна деполяризація на відміну від пейсмекеров відсутня. Швидкість наростання деполяризації велика, висхідна частина ПД дуже крута, але реполяризация протікає уповільнено, і мембрана залишається деполяризованої протягом сотень мілісекунд (див. Рис. 6.3, Б).
Таким чином, тривалість ПД в кардіоміоцитах у багато разів більше, ніж в інших м'язових волокнах. Завдяки цьому всі м'язові волокна передсердь або шлуночків встигають скоротитися до того, як будь-яка з цих волокон почне розслаблятися. Тому фаза реполяризації триває протягом всієї систоли. Під час розвитку ПД збудливість серця, як і інших збудливих тканин, змінюється. Під час деполяризації збудливість серця різко знижується. Це - фаза абсолютної рефрактерності. Причиною її є інактивація натрієвих каналів, що припиняє надходження нових іонів натрію в мембрану. Якщо в скелетної м'язі абсолютна рефрактерність дуже короткочасна, вимірюється десятими частками мілісекунди і закінчується на початку скорочення м'яза, то в серці абсолютна невозбудімості триває весь період систоли. Практично це означає, що якщо під час систоли на серце діє який-небудь подразник, навіть сверхпороговое, то серце на нього не реагує. Тому на відміну від скелетних м'язів серце не здатне до тетанічних скорочень і захищене від занадто швидкого повторного порушення і скорочення. Всі скорочення серцевого м'яза поодинокі. При дуже великій частоті імпульсів збудження серце скорочується не на кожен ПД, а на тільки ті з них, які надходять після закінчення абсолютної рефрактерності.
Під час спадної фази реполяризації, яка збігається з початком розслаблення серцевого м'яза, збудливість серця починає відновлюватися. Це - фаза відносної рефрактерності. Якщо на початку діастоли на серце діє будь-який додатковий подразник, то серце готове відповісти на нього новою хвилею збудження. Позачергове збудження і скорочення серця під дією подразника в період відносної рефрактерності називається екстрасистолою.
Якщо вогнище позачергового збудження знаходиться в синусному вузлі, то це призводить до передчасного виникнення серРіс. 6.4. екстрасистола:
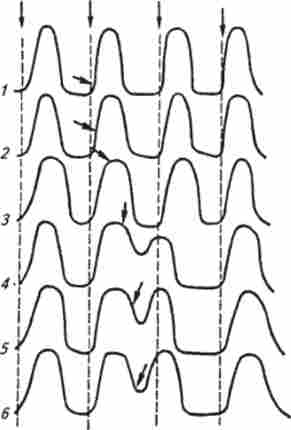
У, 2, 3 - штучне роздратування наносять під час скорочення шлуночків, тому ефекту немає (абсолютна рефрактерна фаза); 4, 5, 6-роздратування наносять під час розслаблення мишіи і викликають появу екстрасистол. амплітуда яких тим більше, чим пізніше в період розслаблення нанесено роздратування. Це пов'язано зі зміною збудливості мишіи, наступаючим після порушення; прямі стрілки і пунктирні лінії вказують момент появи імпульсів в синусовому вузлі, бічні - момент штучного подразнення серця
дечний циклу, при цьому послідовність скорочень передсердь і шлуночків не змінюється. Якщо ж порушення виникає в шлуночках, то після позачергового скорочення (екстрасистоли) з'являється подовжена пауза. Інтервал між екстрасистолою і наступної (черговий) систолой шлуночків називається компенсаторною паузою (Рис. 6.4.).
Компенсаторна пауза пояснюється тим, що екстрасистола, як і будь-яке скорочення серцевого м'яза, супроводжується рефрактерній паузою. Черговий імпульс, що виникає в синусному вузлі, приходить в шлуночки під час абсолютної рефрактерності і не викликає їх скорочення. Нове скорочення настане лише у відповідь на наступний імпульс, коли збудливість міокарда відновиться.
Після відносної рефрактерності в серці настає дуже короткий період підвищеної збудливості - екзальтації, коли серце готове відповісти навіть на подпороговое роздратування.
Провідність. Провідність - властивість серцевого м'яза проводити збудження.
Як уже сказано, імпульс збудження (ПД), виникаючи в пейс- Мекера синусового вузла, поширюється спочатку на передсердя. У передсердях, де дуже невелика кількість які проводять атипових м'язових волокон, порушення поширюється не тільки по ним, але і по робочим кардиомиоцитам. Це пояснює невелику швидкість поширення збудження в передсердях.
Оскільки синусний вузол розташований в правому передсерді, а швидкість передачі ПД невелика, то збудження правого передсердя починається трохи раніше, ніж лівого. Скорочення ж лівого і правого передсердь відбувається одночасно.
Після того як збудження охопить м'язи передсердь, вони скорочуються, а збудження концентрується і затримується в атріовентрикулярному вузлі. Атріовентрикулярна затримка триває до закінчення скорочення передсердь, і тільки після цього збудження переходить на пучок Гіса. Таким чином, біологічне значення атріовентрикулярної затримки полягає в забезпеченні послідовності скорочень передсердь і шлуночків. Одночасне їх скорочення іноді буває при дуже серйозну патологію, коли збудження виникає не в синусному вузлі, а в атріовентрикулярному і поширюється в обидві сторони від атріовентрикулярного вузла - і в передсердя, і в шлуночки. В такому випадку настає різке порушення гемодинаміки в серце.
Механізми атріовентрикулярної затримки не з'ясовані. Можливо, впливає низька амплітуда ПД в клітинах-пейсмекером даного вузла, сильна натрієва інактивація, великий опір міжклітинних контактів.
Далі збудження поширюється по пучку Гіса, ніжок пучка Гіса і волокнам Пуркіньє. Волокна Пуркіньє контактують з скоротливі волокнами міокарда, і збудження передається з провідної системи на робочі м'язи.
Швидкість поширення збудження в серці наступна: від синусового вузла до атріовентрикулярного вузла - 0,5 ... 0,8 м / с; в атріовентрикулярному вузлі - 0,02 ... 0,05; по провідній системі шлуночків - до 4,0; в скорочувальної м'язі шлуночків - 0,4 м / с.
Безпосередній зв'язок провідної системи серця з робочими кардиомиоцитами здійснюється за допомогою численних розгалужень волокон Пуркіньє. Передача сигналів відбувається електричним шляхом з невеликою затримкою. Ця затримка збудження сприяє підсумовування імпульсів, неодночасно надходять по волокнах Пуркіньє, і забезпечує кращу синхронізацію процесу збудження робочого міокарда.
У робочому міокарді є контакти як між торцями, так і бічними поверхнями волокон. Тому збудження від основних стовбурів провідної системи (ніжок пучка Гіса) практично одночасно поширюється на правий і лівий шлуночки, забезпечуючи їх одночасне скорочення.
Напрямок збудження всередині шлуночків різному у тварин різного виду. Так, у собак збудження спочатку виникає на відстані декількох міліметрів від внутрішньої поверхні м'язової стінки, а потім переходить до ендокардит і епікардом. У копитних (у кіз) напрямок поширення збудження в товщі м'язової стінки змінюється багато разів, і безліч волокон в районах ендокарда, епікарда і в глибині стінки активується практично одночасно.
У міжшлуночкової перегородки збудження починається в центральній частині і рухається до верхівки і атріовентрикулярної перегородки, причому верхня частина шлуночків активується пізніше; проте на правій і лівій сторонах міжшлуночкової перегородки збудження виникає одночасно.
Особливості поширення збудження в серці мають значення при аналізі електрокардіограми - записи біострумів серця.
Скорочення. Скорочення - специфічний ознака порушення серцевого м'яза. Як і в інших м'язах, скорочення серцевих м'язових волокон починається після поширення потенціалу дії по поверхні клітинних мембран і є функцією міофібрил. Скорочувальна система міофіб- Рилл представлена чотирма білками - актином, міозином, тро- поніном і тропоміозіном. Скорочення міофібрил серця в принципі не відрізняється від скорочень скелетних м'язів відповідно до теорії ковзання протофібрілл Хакслі.
Суть теорії Хакслі полягає в ковзанні тонких актіно- вих ниток в проміжки між товстими міозіновимі нитками, що призводить до вкорочення саркомера. При розслабленні м'язи Актинові нитки відсуваються назад, займаючи вихідне положення. У механізмі ковзання Актинові ниток має значення кальцій, депонований в саркоплазматичному ретикулуме.
Послідовність електричних і механічних процесів при скороченні серцевих м'язових волокон в даний час представляється в такий спосіб. Потенціал дії, що виник на поверхні мембрани м'язового волокна, по поперечним Т-трубочках, які є впячиваниями зовнішньої мембрани, досягає системи поперечних трубочок, з'єднаних з цистернами саркоплазматичного ретикулума. Порожнини саркоплазматичного ретикулуму не повідомляються ні з Т-трубочками, ні з інтерстиціальної рідиною і заповнені розчином з високим вмістом іонів кальцію. Порожнини Т-трубочок мають такий же склад, що і міжклітинна рідина.
Під час збудження активуються натрієві канали в мембранах Т-трубочок і в міоплазми входять іони натрію і кальцію з міжклітинної рідини. Велика частина входить кальцію не бере участі в скороченні міофібрил, а поповнює його запаси в саркоплазматичному ретикулуме. Під впливом потенціалу дії підвищується проникність мембрани саркоплазматичного ретикулума і іони кальцію виділяються з нього в міоплазми. Іони кальцію зв'язуються з тропонином, що викликає конформаційні зміни в його молекулі. Зрушення тропонин-тропоміозинового стрижня забезпечує взаємодію ниток актину і міозину (нагадаємо, що в розслабленому м'язі актинові волокна прикриті молекулами тропонина і тропомиозина, що утворюють комплекс, що перешкоджає ковзанню протофібрілл).
Після звільнення Актинові ниток від блокування стежка миозинового комплексом міозіновие головки приєднуються до відповідного центру Актинові ниток під кутом 90 °. Потім настає спонтанний поворот головки на 45е, розвивається напруга і відбувається просування актиновой нитки на один крок. Ці процеси здійснюються за рахунок енергії АТФ, причому розпад АТФ каталізує актомиозинового комплексом, що володіє АТФ-азной активністю.
Коли збудження припиняється, вміст іонів кальцію в міоплазми знижується внаслідок роботи кальцієвого насоса і закачування кальцію в саркоплазматичний ретикулум, причому на роботу кальцієвого насоса також витрачається енергія АТФ. В результаті зниження вмісту кальцію в міоплазми стежка міозінових комплекс захищає активні центри актоміозіно- вих ниток. Нитки міозину і актину відновлюють початкове положення, і м'яз розслабляється.
Викладена теорія скорочення серцевого м'яза багато в чому пояснює експериментальні та клінічні спостереження про вплив кальцію і магнію - його антагоніста на роботу серця. Відомо, що при перфузії ізольованого серця розчином, що не містить кальцію, воно зупиняється, а при додаванні кальцію в перфузійний розчин скорочення відновлюються. Відомо також, що серцеві глюкозиди (наприклад, препарати наперстянки) збільшують проникність мембран для кальцію і тим самим відновлюють транспорт кальцію між саркоплазматичним ретикулумом, зовнішньої мембраною і міоплазми.
Узгоджується з теорією м'язового скорочення і сприятливий вплив на серце макроергічних речовин, енергія яких використовується не тільки для механічного скорочення, але і для роботи іонних насосів - кальцієвого і калієво-натрієвого.
Скоротливі властивості серцевого м'яза дещо відрізняються від скелетних. Якщо скелетний м'яз реагує на подразнення відповідно до його силою, то серцевий м'яз підкоряється закону Боудича «все або нічого». Його суть полягає в тому, що на підпорогової роздратування серце не скорочується ( «нічого»), а на порогове подразнення відповідає максимальним скороченням ( «все»), і збільшення сили подразника не призводить до збільшення сили скорочення.
У скелетних м'язах закону «все або нічого» підкоряються окремі м'язові волокна. Справа в тому, що потенціал дії викликає звільнення кальцію з саркоплазматичного ретикулума рівномірно по всій довжині волокна, тому воно скорочується повністю. Але в скелетної м'язі є волокна з різним ступенем збудливості, тому при слабкому подразненні скорочуються не всі волокна і сумарне скорочення виявляється невеликим. У серцевої ж м'язі волокна робочого, т. Е. Скорочувального, міокарда з'єднані міжклітинних контактів
(Виростами плазматичних мембран), що сприяє практично одночасному поширенню потенціалу дії по всій м'язі, і вона збуджується і скорочується як єдиний орган, будучи функціональним сінцітіем.
Закон Боудича є скоріше правилом з певними обмеженнями. При підпороговому подразненні скорочення, дійсно, не виникає, але в цей час починається активація натрієвих каналів і підвищується збудливість міокардіоцитів. Виникаючі місцеві потенціали можуть підсумовуватися і викликати поширюється потенціал дії. З іншого боку, сила скорочення серця, як добре відомо, непостійна і може змінюватися в різних умовах життя.
Інша характерна риса серцевого м'яза полягає в тому, що сила скорочення серця залежить від ступеня розтягування м'язових волокон під час діастоли, коли порожнини заповнюються кров'ю. Це - закон Франка - Старлінг. Зазначена закономірність пояснюється тим, що при розтягуванні серця кров'ю під час діастоли Актинові нитки кілька витягуються з проміжків між міозіновимі, і при подальшому скороченні зростає число генеруючих силу поперечних містків. Крім того, при розтягуванні серцевого м'яза в ній підвищується опір пружних елементів, і під час скорочення вони грають роль «пружини», збільшуючи силу скорочення.
Особливо важливе значення закон Франка - Старлинга має під час посиленої роботи серця, коли зростає обсяг крові, що надходить в нього під час діастоли. Збільшення сили скорочення призводить до того, що вся кров викидається під час систоли шлуночків в артеріальні судини, інакше після кожного скорочення в серці залишалася б значна порція крові. При відсутності великого навантаження і невеликому обсязі кровотоку сила скорочення серця помірна. Таким чином серце здатне регулювати в певних межах силу скорочення в залежності від обсягу кровотоку.