| Головна |
| «« | ЗМІСТ |
|---|
Під час збудження нейрон генерує так званий потенціал дії - різкий стрибок мембранного потенціалу в позитивну сторону. Потенціал дії (ПД, спайк, імпульс) виникає на мембранах збудливих клітин (нервових і м'язових, а також деяких рецепторних і секреторних клітин). Виникненню ПД зазвичай передує деполяризация клітинної мембрани. Для цього необхідний Деполяризуючий ток (як правило, це струм, створюваний входять позитивними зарядами). Цей струм може виникати при електричному впливі на клітку (якщо, наприклад, порушується сусідня тісно пов'язана клітина), але найчастіше при дії на клітину спеціальних збуджуючих речовин, про які мова піде пізніше.
Якщо величина деполяризующего струму менше порогового значення, то виникає лише пасивна деполяризація мембрани, нераснространяю- щееся збудження, зване локальний відповідь. Якщо деполяризація досягає порогового рівня, то розвивається ПД. Після досягнення порогового потенціалу подальша деполяризація наростає лавиноподібно, потенціал стає все менш негативним і в якийсь момент навіть позитивним (так званий овершут).
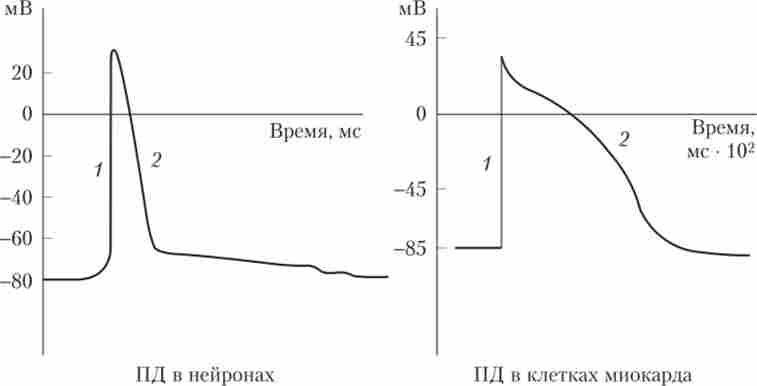
Мал. 3.6. Види потенціалів дії в клітинах різного типу:
1 - деполяризация; 2 - реполяризация
У нормі можна зареєструвати або максимальний електричний відповідь мембрани (IIД), або дуже невеликий локальний відповідь, але не якісь небудь проміжні за величиною реакції. Тому кажуть, що ПД підкоряється закону «все або нічого», Т. Е. Амплітуда ПД завжди максимальна і не залежить від сили подразника (рис. 3.7).
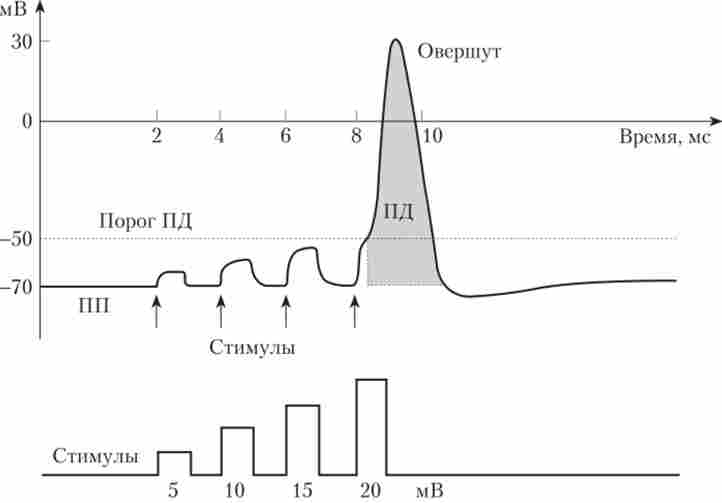
Мал. 3.7. Розвиток потенціалу дії при подразненні нейрона збільшуються за амплітудою електричними стимулами1
Ще одна характерна особливість ПД - швидка реполяризация до рівня спокою після досягнення максимуму. Тривалість ПД може бути різною: в деяких нервових волокнах вона становить мілісекунди, а в серцевому м'язі - близько 0,5 с. При зменшенні інтервалу між двома ПД величина другого з них стає все менше і менше. Якщо повторний стимул (Деполяризуючий ток) подається дуже швидко після закінчення першого ПД, то другий ПД не розвивається. Цей період стану нейрона називається період рефрактерності (див. нижче).
З 1936 р улюбленим об'єктом для дослідження функцій збудливих мембран були так звані гігантські аксони кальмара. Завдяки їх великому діаметру (до 1 мм) на них легко працювати, тому перші великі відкриття були зроблені саме на цих аксонах. У 1949 р А. Ходжкин і Б. Кац висунули так звані натрієву гіпотезу природи ПД, згідно з якою фаза підйому і овершут ПД обумовлені тимчасовим підвищенням проникності мембрани для Na+ і входом цього іона в клітку.
На користь натрієвої гіпотези говорить наступне:
Показано, що при дії деполяризующего струму на клітину спостерігається спочатку дуже швидкий вхідний струм, а через 1-2 мс він змінюється повільніше розвиваються виходять струмом. Перший визначається вхідними іонами Na+ і відповідає за швидку деполяризацію, другий визначається виходять іонами К+ і відповідає за реполяризацию.
Вхідний натрієвий струм. При порушенні мембрани іони Na+ входять через спеціальні канали в мембрані, вибірково проникні для цих іонів. иа+-канали активуються (т. е. відкриваються і пропускають іони) у відповідь на деполяризацію, при цьому вони вибірково пропускають переважно іони Li+ і Na+. Така властивість обумовлено особливою молекулярною структурою каналів, завдяки якій вони грають роль селективних фільтрів. Білкові молекули, що формують канали, можуть змінювати свою конформацію (структурну організацію) під дією змін МП, в результаті чого канали відкриваються (активуються), закриваються і инактивируются. Передбачається ворітної механізм роботи Ка+-каналу. Блокатором цих каналів (речовиною, що зв'язуються з молекулами каналів і блокуючим їх роботу) є тетродоток- син - отрута, виділений з внутрішніх органів риби іглобрюха. Яд як би затикає канал. У лабораторних експериментах його використовують для блокади передачі нервових імпульсів.
Цикл Ходжкіна. При деполяризації число відкритих №+-каналів збільшується. збільшення Иа+-провідності лежить в основі тих само- посилюються процесів, через які ПД підкоряється закону «все або нічого». При дії деполяризующего стимулу спочатку відбувається відкривання лише невеликого числа Иа+-каналів. Через них деяку кількість іонів Na+ увійде в клітку. Вони принесуть позитивні заряди і додатково деполярізуют клітку. Це призведе до відкривання додаткових Иа+-каналів і т. д. Такий замкнутий круг, що зв'язує МП і Иа+-провідність, називається циклом Ходжкіна (приклад позитивного зворотного зв'язку).
Вихід калієвий струм відповідає за реполяризацию клітини. У міру того як МП стає все більш позитивним, збільшується електрорушійна сила (ЕРС), що діє на іони К+, т. е. іони К+ прагнуть покинути клітку. Канали, що відповідають за цей струм, також активуються при деполяризації. Але вони відкриваються повільніше, ніж Иа+-канали, тому ПД встигає розвинутися без перешкод. відмінність Na+- і К+-каналів в тому, що при деполяризації Иа+-канали швидко відкриваються і швидко інактивуються йод дією деполяризації, далі Иа+-канали можуть знову відкритися, тільки коли мембрана буде знову реполярізована до вихідного рівня МП. А К+-канали відкриваються повільно і не інактивуються при деполяризації, а закриваються тільки тоді, коли мембрана реіолярізуется до вихідних значень.
Звідси виникає такий стан збудливою клітини, як рефрактер- ність - стан тимчасової невозбудимости мембрани клітини. Під час ПД відбуваються два процеси, в результаті яких нейрон не здатний якийсь час генерувати наступний потенціал дії. По-перше, інактивація натрієвих каналів досягає найбільшого рівня під час фази спаду ПД, і на її усунення необхідно кілька мілісекунд. По-друге, завдяки активації калієвих каналів проникність для цих іонів дуже висока на спаді ПД і поступово повертається до рівня спокою. При таких умовах для деполяризації треба було б дуже велике збільшення проникності для іонів натрію.
Обидва перерахованих чинники призводять мембрану клітини в стан абсолютної рефрактерності - повної невозбудимости, коли навіть надпорогові стимули не можуть викликати повторного відповіді у вигляді ПД. Після абсолютної рефрактерності настає період відносної рефрактерності, в ході якого поріг поступово знижується до нормального рівня, натрієві канали відновлюються з інактивованого стану, а калієві канали закриваються (рис. 3.8). Під час відносної рефрактерності Деполяризуючий сигнал досить великий амплітуди може активувати таке число Иа+-каналів, щоб, незважаючи на велику кількість ще відкритих До+-каналів, виник ПД. Однак внаслідок високої До+-провідності і значною залишковою Na+- інактивації МП в точці максимуму не буде настільки близький до?Na, а отже, амплітуда ПД буде менше.
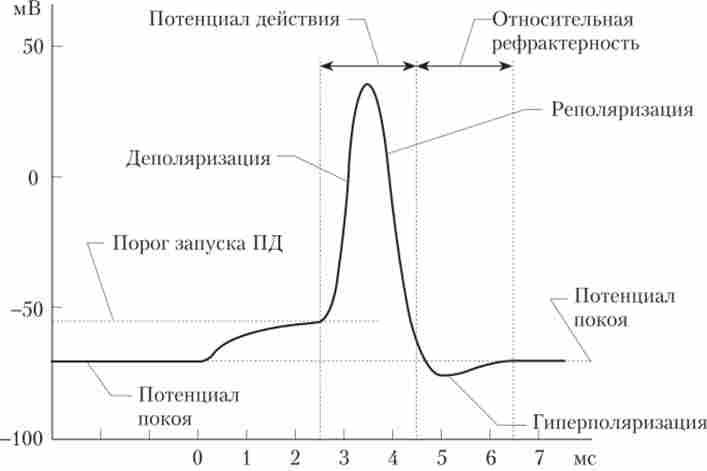
Мал. 3.8. Етан розвитку потенціалу дії і рефракгерность1
Важливо пам'ятати, що активний транспорт іонів через мембрану (наприклад, Na+/ K+-Hacoc) не відповідає безпосередньо за фази деполяризації і реполяризації ПД, але необхідний для підтримки іонних градієнтів, які відповідальні за виникнення мембранних струмів.
Існують також і інші Електровозбудімость канали. У багатьох збудливих клітинах є потенціалзалежні Са2+-канали. Однак Са2"- струми не вносять істотного внеску в розвиток ПД. Са2+ в клітці, як правило, виступає в якості посередника, що запускає багато внутрішньоклітинні процеси, секрецію і т. д.