| Головна |
| «« | ЗМІСТ | »» |
|---|
Сучасні уявлення про молекулярний механізм кроссінго- віра в основному склалися в 1960-і рр. При цьому з урахуванням особливостей молекулярної структури ДНК як носія генетичної інформації більш детально розроблена гіпотеза «розрив-возз'єднання».
Найбільшу популярність придбала модель Р. Холлідей (рис. 6.7). Рекомбінація відбувається тільки між двома хроматидами. Ще дві хроматиди залишаються інтактними, однак при розгляді кінцевого результату - розщеплення в тетрадах - їх також необхідно врахувати. АВС і abc - три тісно зчеплених маркера, доля яких простежується протягом усього процесу рекомбінації. Стрілки, що показують антіпарал- лельного ланцюга ДНК. Для даної схеми (рис. 6.7) дуже істотний облік полярності ланцюгів. Необхідно мати на увазі, що на схемі на стадіях а-г (Ґ) показані тільки дві хроматиди з чотирьох - дві двунітевие молекули ДНК, встулающіе в рекомбінацію. Дві інші хроматиди, які не вступають в рекомбінацію, не показані. На стадії д (> д ') - розщеплення в тетрадах з урахуванням всіх чотирьох хроматид, як показаних, так і не показаних на попередніх стадіях
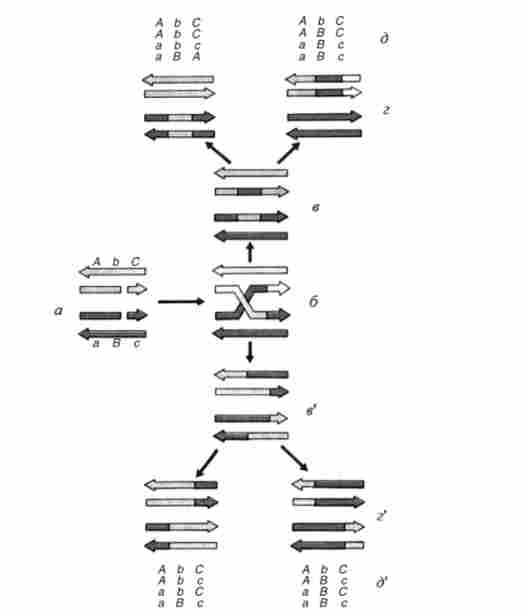
Мал. 6.7. Схема кроссінговсра, запропонована Р. Холлідеєм: а-д стадії
Весь процес ініціюють два однониткових розриву в нитках однакової полярності. Вони зафіксовані в гомологічних точках, однак це не має принципового значення. Розриви можуть і не бути строго гомологічних. На першому етапі молекули ДНК, що вступають в рекомбінацію, утворюють гібридні ділянки - так звані гетеродуплекси, в яких одна ланцюг походить від однієї молекули, а інша - від іншої. це напів хіазма. На наступному етапі в точці перехрещення нитки розриваються. При цьому рвуться або нитки, в яких були первинні розриви (рис. 6.7, д), або дві інші нитки. Передбачається, що обидва типи розривів різновірогідні (рис. 6.7, в).
Таким чином, виходять або дві нерекомбінантні по фланговим маркерами молекули (А-С і а-з), Що несуть гібридний ділянку - зону гетеродуплекса в районі середнього маркера В / b (Рис. 6.7, в), або дві молекули, рекомбінантні по фланговим маркерами, теж гетеродуплексние в районі середнього маркера (рис. 6.7, в ').
Оскільки, згідно з положенням Дж. Уотсона і Ф. Кріка, мутації - це зміни чергування нуклеотидів в ДНК, аллели одного гена, зокрема В / b, розрізняються за складом нуклеотидів (як мінімум по одній парі підстав). Тоді в ділянці гетеродуплекса повинна утворитися зона локального неснаріванія підстав. Ці ділянки «дізнаються» спеціальні ферменти репарації, що забезпечують структурну стабільність ДНК. Вони усувають ту чи іншу неспареними підставу і замінюють його на комплементарное. У такому процесі корекції з однаковою ймовірністю матрицею служить та чи інша нитка гетеродуплскса (рис. 6.7, г, ґ). Однак в кожному конкретному випадку корекція поширюється уздовж хроматиди? і в якості матриці використовується одна і та ж одиночна нитка гетеродуплекса ДНК. Якщо в зону гетеродуплекса потрапляє кілька маркерів, то це повинно привести до коконверсіі.
В результаті розщеплення в тетрадах можна розглядати як результат молекулярного механізму кросинговеру (рис. 6.7, д с) '). Якщо взяти до уваги ще дві хроматиди, які не вступали в рекомбінацію, буде ясно, що схема з однаковою ймовірністю допускає появу зошити з конверсією 3: 1 і 1: 3 як з кросинговером за середнім маркера (рис. 6.7, д так і без нього по фланговим маркерами (рис. 6.7, д т. е. згідно з цією схемою кореляція конверсії і кросинговеру становить 50%.
Якщо уявити, що корекція на стадії гіг ' відбудеться тільки в одній хроматиді, а в іншій - не відбудеться, то буде спостерігатися постмей- отіческос розщеплення (зь : 5В або 5b : 3В), зустрічається в октадах у нейроспори. Його відзначають і в тетрадах дріжджів. У цьому випадку деякі гаплоїдні колонії, вирощені з аскоснор однієї тетради, будуть представлені половинками різного генотипу: В і Ь. Таке явище дійсно спостерігається у дріжджів.
Істотний момент в розглянутій схемі - необхідність додаткового синтезу ДНК в процесі рекомбінації, зокрема при репарації (корекції) гетеродуплексов. Дійсно, І. Хотта і X. Штерн виявили у рослин (лілії) і у тварин (миші) в пахітене мейозу невеликий синтез ДНК репаративного типу, який доповнює основну реплікацію ДНК в премейотічної 5-фазі. У тих же об'єктів на стадії зиготи-пахітени показано підвищення активності ферменту, що виробляє однонітевиє розриви в ДНК, а також посилений синтез білка, що дестабілізує подвійну спіраль ДНК. Все це необхідно для рекомбінації.
На особливу увагу в схемі Р. Холлідей заслуговує спосіб утворення гетеродуплексов. Показані послідовні стадії утворення підлозі хіазми (А-В), Яка потім може видозмінюватися шляхом міграції вздовж кон'югується хроматид (молекул ДНК) на стадіях в, г (Рис. 6.8). Цей процес отримав назву міграції гілок полухіазми. Зона перекидання рухається подібно застібці «блискавка», подовжуючи ділянки гетеродуплексов. При цьому молекули ДНК повинні обертатися навколо своїх осей назустріч один одному. Експерименти з об'ємними молекулярними моделями показують, що це можливо. Результати зошитового аналізу у дріжджів, гетерозиготних за двома маркерами, між якими відомо відстань, виражене в кількості пар нуклеотидів, показують, що зона гібридної ДНК може поширюватися на ділянки довжиною близько 1 000 п. Н. Про це судять по здатності до коконвсрсіі мутацій, розташованих на такій відстані один від одного.
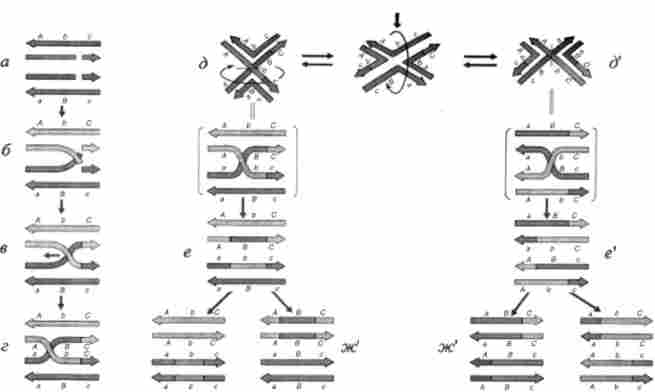
Мал. 6.8. Освіта підлозі хіазми: a-в- мшрація гілок; г- ізомеризація полухіазми; д, д ' - різні результати дозволу полухіазми в залежності від характеру розривів (горизонтальна і вертікатьная лінії в зоні перекидання гібридної ДНК на стадії д) еу е ж, ек? - корекції гетеродуплексов. У дужках - варіанти зображення ізомерів полухіазми (г). Демонструються тільки дві хроматину з чотирьох - ті, що вступають в рекомбінацію
Модель Р. Холлідей деталізована і в тій частині, яка пов'язана з дозволом полухіазми двома типами розривів, що призводять потім до конверсії без кросинговеру в одному варіанті і до конверсії з кроссінго- Вером - в іншому. Якщо фігуру, утворену внаслідок міграції гілок полухіазми (рис. 6.8, г), зобразити трохи інакше (рис. 6.8, д), то вона допускає обертання пов'язаних полухіазмой молекул відносно один одного (рис. 6.8, ду д '). Цей процес називається изомеризацией полухіазми. Розрив в точці перехрещення одиночних ниток призводить до конверсії без кросинговеру (рис. 6.8, д) або до конверсії з кросинговером по фланговим маркерами (рис. 6.8, д '). Характерні фігури, відповідні стадіях рекомбінації, були виявлені при електронно-мікроскопічному вивченні рекомбінації молекул ДНК плазмід в бактеріальних клітинах.
Незважаючи на великі успіхи в розумінні механізму гомологичной рекомбінації (загальною рекомбінації), залишається ще багато питань. Основні етапи рекомбінації у всіх організмів, мабуть, подібні, проте не зовсім зрозумілі відмінності в цьому процесі, які обумовлені різницею в організації генетичного матеріалу бактерій і еукаріот з їх нуклеосомної структурою хроматину. Неясно, як і роль СК в процесі рекомбінації, які ініціюють події в освіті полухіазми і т. Д. Чи не вивчені деталі митотичного кросинговеру. Відомо, що зони гібридної ДНК при митотичної конверсії довше, ніж при мейоті- чеський, однак відсутність регулярної кон'югації хромосом в мітозі ускладнює розуміння всього процесу.
Незважаючи на ряд невирішених питань, розробка молекулярного механізму рекомбінації є яскравим прикладом синтезу формально-генетичного і молекулярно-біологічного підходів в дослідженнях одного з найважливіших біологічних процесів.