| Головна |
| «« | ЗМІСТ | »» |
|---|
Розпад глікогену і гліколіз швидко активуються вже в перші секунди інтенсивних навантажень. Якщо навантаження триває, починає рости внесок гліколізу в ресинтез АТФ.
Розпад глікогену до глюкозо-6-фосфату і потім до лактату проходить через ряд стадій (див. Гл. 4). При цьому продукти гідролізу АТФ і РСГ (АДФ. АМФ, ІМФ, NH3 і Р |), утворені в результаті м'язової роботи, стимулюють розпад глікогену. Тим самим прискорюється підживлення анаеробної роботи м'язів.
Гліколіз здійснюється через більшу кількість стадій, ніж гідроліз РСГ. Тому освіту АТФ при анаеробному гліколізі йде повільніше, ніж при гідролізі РСГ, але набагато швидше, ніж при окисного фосфорилювання.
Довгий час припускали, що РСГ - єдине біопаливо, яке забезпечує початкові етапи м'язової роботи до тих пір, поки концентрація РСГ не стане низькою і не підключиться утилізація глікогену.
Насправді катаболізм глікогену може служити основою для енергозабезпечення максимальних зусиль, які тривають від 20 с до приблизно 2 хв. За цей час витрачається істотна частка м'язовогоглікогену (близько 25 ммоль глюкозних одиниць / кг).
Виявляється, що кількість АТФ, утвореної за рахунок глюкози з м'язовогоглікогену, набагато вище, ніж за рахунок гідролізу всього запасу РСГ. У спринті запас глікогену швидко виснажується з утворенням лактату. Частина цього лактату йде з м'язів в кров.
Фосфагеновая система може забезпечити вихід потужності нижчий, ніж забезпечує гідроліз глікогену. Тому максимальна швидкість гідролізу при виконанні того чи іншого виду роботи не може підтримуватися більш декількох секунд.
Скелетні м'язи людини складаються принаймні з двох типів метаболічно і функціонально різних волокон.
Тип I волокон характеризується повільним скороченням, здатністю до тривалої роботи без втоми, відносно низькою вихідною потужністю і переважним анаеробним ресинтезом АТФ при скороченні.
Тип II волокон характеризується відносно швидкою сократимостью, швидкою втомою, високою вихідною потужністю і переважним анаеробним ресинтезом АТФ.
Комбіноване взаємодія волокон різного типу забезпечує виконання різноманітних м'язових навантажень.
Додатковим шляхом регенерації АТФ, коли запаси АТФ і РСГ виснажуються, є міокіназная реакція. У міокіназной реакції взаємодіють дві молекули АДФ з утворенням однієї молекули АТФ і однієї молекули аде- нозілмонофосфата АМФ:
Міокіназная реакція каталізується ферментом міокіназа. Ця реакція стає важливою під час високоінтенсивних рухів, коли поставка енергії через АТФ вкрай обмежена. АМФ є аллостеричним фактором для багатьох ферментів реакцій гліколізу і розщеплення глікогену.
Окислювальний метаболізм також дає внесок в ресинтез АТФ за 10 з спринту на 100 м. Хоча це всього 10% загального ресинтезу АТФ, вони дуже важливі для збільшення часу швидкого бігу.
При метаболічному кризі під час великих навантажень великого значення набуває розпад АМФ до аденіну і і нозі л монофосфата (ІМФ).
Зменшення вмісту АМФ в м'язах може здатися контрпродуктивним. Але цей процес починається лише при малих значеннях відношення концентрацій АТФ / АДФ, запобігаючи надмірне накопичення АДФ і АМФ. Це забезпечує можливість протікання аденілатціклазной реакції і роботи м'язів шляхом підвищення відносини АТФ / АДФ.
Концентрації АТФ. АДФ і АМФ можуть бути використані для розрахунку енергетичного заряду Е ,, (запасу) клітини по Аткінсон. Ця величина вимірюється ступенем фосфорилювання Аден л нуклеотидного запасу клітини і розраховується за формулою
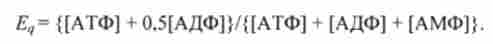
Енергетичний заряд є хорошим індикатором енергетичного статусу клітини. Наприклад, якщо енергетичний заряд дорівнює 1,0, це означає, що весь фосфор знаходиться у формі АТФ і клітина максимально готова до роботи. В іншому крайньому випадку, якщо енергетичний заряд дорівнює нулю, це означає, що АТФ повністю гідролізовані до АМФ.
Розглянуті випадки теоретичні. Насправді запас аденілнукле- отиде не буває повністю в формі АТФ і його рівень не опускається нижче значення 0,6.
У нормі відпочинку енергетичний заряд знаходиться в межах значень 0,90- 0,95. При хвороби він може знижуватися до 0,85, при роботі на витривалість (довгі дистанції) - до 0,7. Якщо енергетичний заряд стає нижче значення 0,7, коли м'язи починають втомлюватися, розпад АТФ відзначено зниження продуктами його розпаду АДФ і АМФ.
Навіть в спринті на 100 м бігуни можуть призвести до затримки на фінішній третини дистанції. Починає позначатися втома м'язів. У спринтерів високого класу випробування на велоергометрі показують, що фаза прискорення - максимум потужності - знаходиться в межах від 3 до 4 с, після чого потужність падає. На доріжці за цей час спортсмен долає приблизно третину дистанції. Після цього швидкість падає.
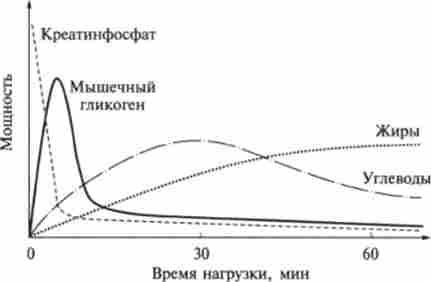
Мал. 13.11. Механізми енергозабезпечення м'язів при різних навантаженнях
Наступ втоми залежить від багатьох факторів, але один з головних - зниження анаеробної продукції АТФ і накопичення АДФ в результаті розпаду РСГ. На втому впливають також накопичення метаболітів і зміна перенесення кальцію.
Під час відпочинку після навантажень йде ресинтез фосфокреатину відповідно до реакцією
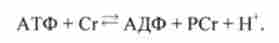
Чим вище були навантаження, тим більше часу потрібно для відновлення, так як реакція йде з обмеженою швидкістю.
Механізми енергозабезпечення м'язів при різних навантаженнях і відповідний вихід потужності представлені на рис. 13.11.
Питання до розд. 13.3